![]() Az agy egyik legfőbb feladata az állandóan változó környezeti ingerekre való válaszreakciók kialakítása, továbbá az azokból kiszűrt, számunkra fontos információk emléknyomok formájában történő elraktározása. Ezen folyamatokhoz szükséges az agy működésének folyamatos finomhangolása: más-más állapotra van szükség egy félelmi, egy romantikus, egy vicces vagy éppen egy alvási állapotban. A közelmúltban a különböző éberségi és alvási állapotokat meghatározó, alapvető fontosságú hálózatot fedeztünk fel, mely eredmények utat nyithatnak az alvászavarok kezelése és radikálisan új típusú altatók fejlesztése felé. A felfedezést leíró cikk az idegtudomány legrangosabb lapjában, a Nature Neuroscience-ben jelent meg.
Az agy egyik legfőbb feladata az állandóan változó környezeti ingerekre való válaszreakciók kialakítása, továbbá az azokból kiszűrt, számunkra fontos információk emléknyomok formájában történő elraktározása. Ezen folyamatokhoz szükséges az agy működésének folyamatos finomhangolása: más-más állapotra van szükség egy félelmi, egy romantikus, egy vicces vagy éppen egy alvási állapotban. A közelmúltban a különböző éberségi és alvási állapotokat meghatározó, alapvető fontosságú hálózatot fedeztünk fel, mely eredmények utat nyithatnak az alvászavarok kezelése és radikálisan új típusú altatók fejlesztése felé. A felfedezést leíró cikk az idegtudomány legrangosabb lapjában, a Nature Neuroscience-ben jelent meg.
Mindenki tapasztalta már, hogy egy váratlan esemény következtében felfokozódik a lelkiállapota (ébersége), öröm vagy félelem, ijedség lesz úrrá rajta, mely kihat nemcsak a következő néhány órára, de akár az éjszakai alvására is. Rosszabb esetekben akár tartós alvászavar is kialakulhat. Hiába készül fel valaki alaposan egy vizsgára vagy egy állás-interjúra, vesz át minden lehetséges kérdést több tucatszor, a teljesítés és számonkérés idején leblokkolhat, túlzott izgalmi állapotában „még a saját nevét is elfelejti”. Az éberségnek, izgatottságnak ezt a fokozódását az agy állapotának hirtelen megváltozása idézi elő. Az hogy, hogyan viszonyulunk a dolgokhoz – izgatottan, motiváltan, türelmetlenül, vagy éppen unottan, kókadtan, álmosan, számos agyi rendszer összehangolt működésének eredménye. E rendszerek alapvetően meghatározzák, hogy egy adott információt, hogyan dolgozunk fel és raktározunk el. De van-e egy olyan karmester az agyban, ami minden érzelemről, indulatról tudomást szerez és független attól, hogy pozitív vagy negatív az élmény, beállítja az izgatottság/éberség szintjét? Jelzi számunkra a dolog fontosságát? Összeköti-e valamilyen agyi rendszer az izgatottság kialakítását és az alvásból való felébredést? Létezik erre egy külön rendszer az agyban?Az MTA Kísérleti Orvostudományi Kutatóintézet és az MTA Természettudományi Kutatóközpont közös kutatóprogramjában több kutatótársunkkal, akik közül kiemelendő Komlósi Gergely, egy olyan idegsejt hálózatot fedeztünk fel, amely éppen ezt a feladatot látja el: beálltja az adott feladat teljesítéséhez szükséges állapotot, és ez által fontos szerepe van az alvás és ébrenlét szabályozásában is.
Különleges középvonali talamikus sejtek az éberség hátterében
A külvilágból, illetve a saját testünkből érkező információk az agy ősibb, agytörzsi területeinek közvetítése révén kapcsolódnak át a talamuszban (BOX 1), és onnan továbbítódnak a legfelsőbb feldolgozói régióba, az agykéreg felé.
A talamusz az agy közepén elhelyezkedő több, különböző funkcióra specializálódott magból álló agyterület. Az agykéreg gyors, specifikus információt szinte kizárólag a talamuszon keresztül kap, így a talamusz legfőbb feladata az agykéreg működésének hatékony kontrollja. Minden agykéregi terület oda-vissza kapcsolatban áll a talamusszal és e kapcsolatok nélkül az agykéreg nem tudja betölteni funkcióját.
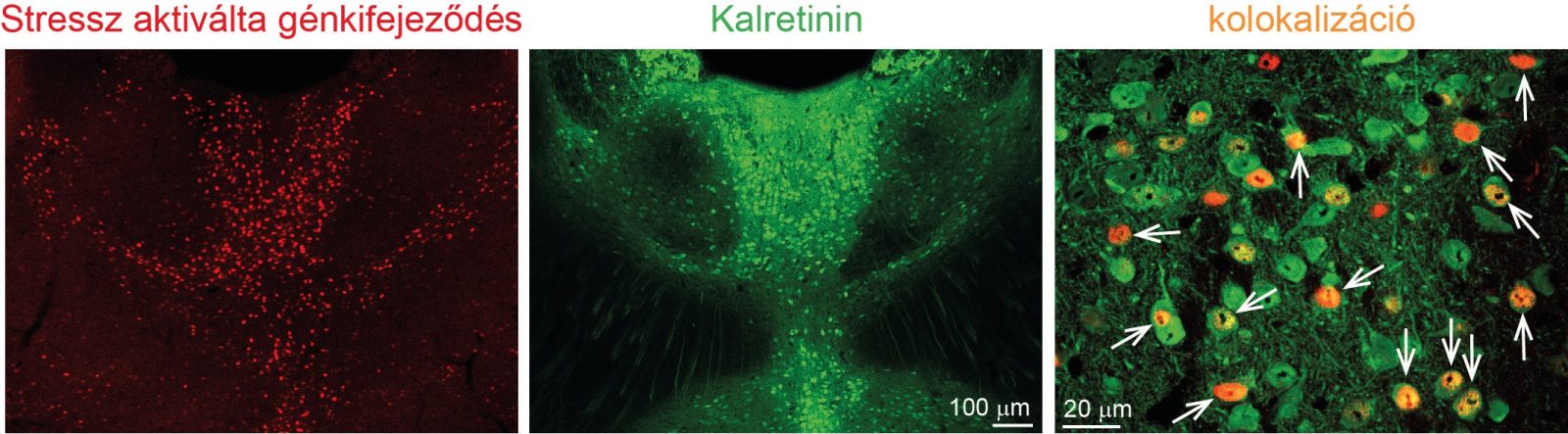
A múlt század 40-es éveiben a New Yorkban dolgozó Moruzzi és Mogun több kortársával már rámutatott arra, hogy a középvonali talamikus területek (KT) különleges szerepet játszhatnak az éberség kialakításában. Feltételezéseik szerint a KT arra specializálódott, hogy összegyűjtse az agytörzsi eredetű serkentő szignálokat és továbbítja a teljes agykéreg területére. Ennek következménye egy, a teljes agyra kiható ébresztés, az agy állapotának gyors megváltozása. Ezzel a serkentő pályarendszerrel az ún. agytörzsi felszálló aktiváló rendszerrel találkozhattunk középiskolai tanulmányaink során is. Az azóta eltelt évtizedekben azonban számos tanulmány megcáfolta a KT ébresztésben betöltött szerepét. Az alapvető probléma az volt, hogy évtizedeken keresztül nem voltak meg a megfelelő módszerek arra, hogy ezen sejtek aktivitását célzottan vizsgálják, működésüket szelektíven befolyásolják és ennek hatását az éberségre precízen mérni tudják. Hét évvel ezelőtt egy váratlan felfedezés révén elszántuk magukat arra, hogy megvizsgáljuk és megpróbáljuk feloldani az ellentmondásokat. Észrevettük ugyanis, hogy az általunk használt kísérleti állat, az egér KT-jának idegsejtjei igen érzékenyen reagálnak minden helyzetre, ami eltér a megszokottól, legyen az akár pozitív, akár negatív a kisegér számára. Ezen sejtek közös tulajdonsága volt, hogy egy kalciumkötő fehérjét, a kalretinin-t (KR, BOX 2) tartalmazták.
A kalciumkötő fehérjék alapvetően befolyásolják a sejtek működését, azáltal, hogy membrán serkentése során beáramló, vagy a sejtszervecskékből kikerülő kalciumot megkötik, így meghatározzák a kalciumhoz köthető élettani folyamatok sebességét. Különböző sejtek, különböző típusú kalciumkötő fehérjét fejezhetnek ki, ami lehetővé teszi ezek sejtek szelektív vizsgálatát.
E tulajdonság alapján jól elkülöníthetők voltak mind anatómiailag, mind pedig élettanilag a körülötte elhelyezkedő, KR-t nem termelő talamikus idegsejtektől (1. ábra). Így rögtön felmerült a lehetőség e sejtek szelektív vizsgálatára és feltettük a kérdést, hogy ezek a sejtek képesek lehetnek-e arra, hogy befolyásolják az agy egyes (éberségi) állapotát. De hogyan vizsgálható egy adott sejttípus? Minden agyterület nagyon sokféle idegsejtet tartalmaz általában egymással elkeveredve. Hogyan választható csak az egyik sejttípus egy kevert populációból? Erre a célra fejlesztették ki az elmúlt 10 évben a genetikai alapú, sejt-szeletkív jelölési technikákat, melyek transzgénikus egerek és kódoló DNS szekvenciát, plazmidot, hordozó virális vektorok felhasználására épülnek (BOX 3).
A sejt-szelektív genetikai alapú technikák során egy adott sejtcsoportba jutattunk egy adott gént. A gén által termelt fehérje nem zavarja a sejt normális működését, azonban lehetőséget nyújt arra, hogy a sokféle sejttípust tartalmazó szövetben adott sejteket szelektíven megjelenítsük (szerkezettani vizsgálatok), működésüket szelektíven vizsgáljuk és befolyásoljuk (élettani vizsgálatok), valamint detektáljuk a megváltoztatott működés hatását a viselkedésre (viselkedéstani vizsgálatok). A módszer lelke egy bakteriális DNS hasító enzim (CRE-rekombináz), ami csak egy jól meghatározott, emlősökben nem előforduló bázis sorrendnél hasítja a DNS-t. Ha készítünk egy virális plazmidot, amibe egy adott fehérjét (pl. egy fényesen világító fluoreszcens fehérjét) kódoló DNS szakasz két végét ezen bázis sorrend „börtönébe” zárjuk, erről a DNS-ről nem tud fehérje átíródni, mikor ezt a plazmidot a szövetbe juttatjuk és az ott lévő sejtek felveszik. Azonban, ha a plazmid találkozik a CRE-rekombináz enzimmel, ez a molekuláris olló elhasítja ezeken a pontokon a DNS szálat és megindulhat a génátírás, esetünkben elkezdhet termelődni a fluoreszcens fehérje. Most már csak azt kell megoldani, hogy CRE-rekombináz enzim csak egy adott proteint (esetünkben a kalretinint) termelő sejtben forduljon elő a többiben nem. Ez pedig a transzgenikus technikákkal jelenleg könnyen megoldható. Tehát, jelen esetben, ha van egy egértörzsünk, amibe csak a kalretinin tartalmú sejtekben fejeznek ki CRE-rekombinázt és ezen állatok talamuszába olyan plazmidot jutattunk, ami a „börtönbe zárt” fluoreszcens fehérjét tartalmazza, akkor csak és kizárólag a kalretinin tartalmú talamikus sejtekben történik meg a fluoreszcens fehérje átírása és, először a világon, rácsodálkozhatunk ezen sejtek nyúlvány rendszerére.
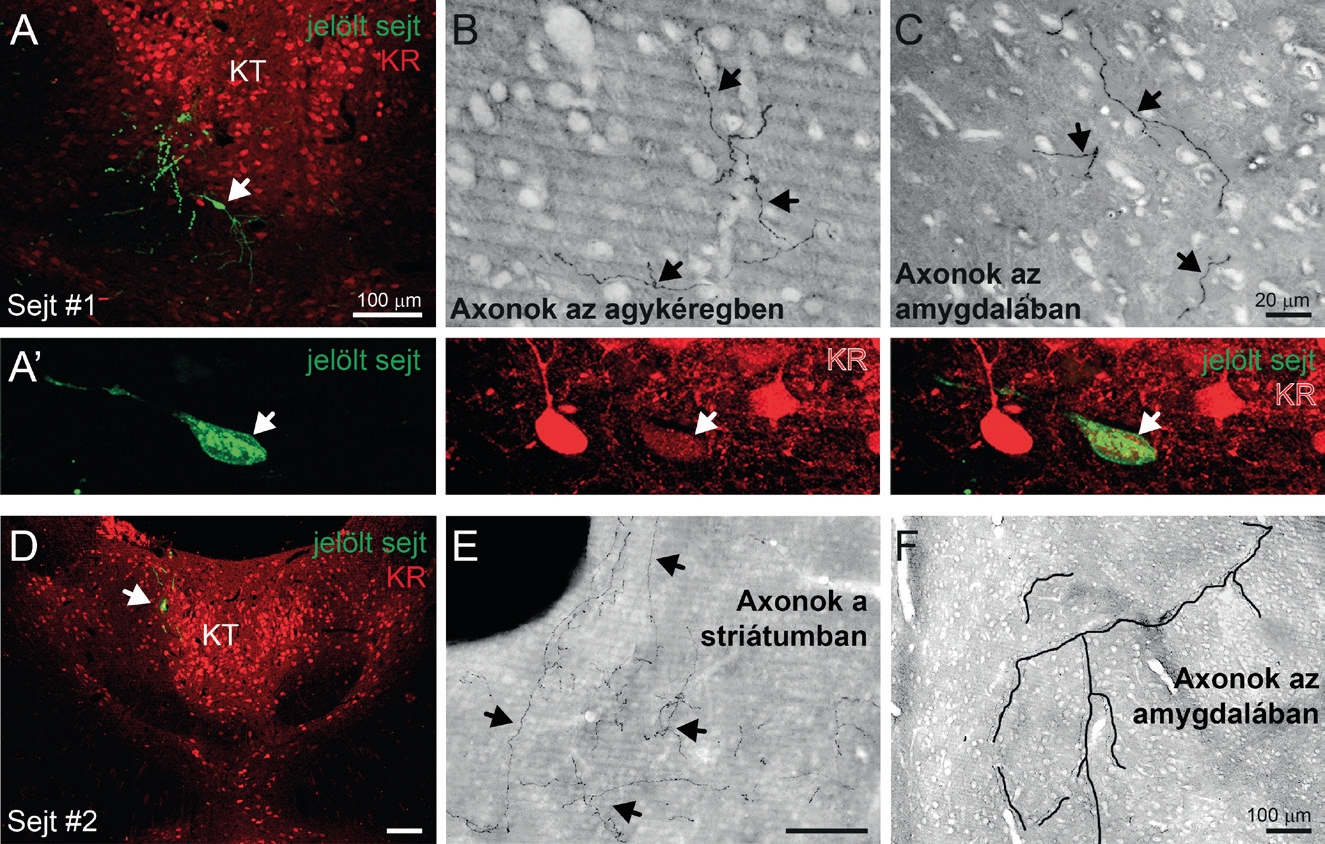
Kísérleteink során a zöld fluoreszcens fehérjét (green fluorescent protein, GFP) kódoló plazmidot injektálunk a KR-Cre egér KT-jába. Így ezen KR+KT sejtek dendritfája és teljes axonális hálózata zölden világított az adott hullámhosszúságú fény megvilágítására. Ahhoz, hogy egy idegsejt-csoport képes legyen az egész agy állapotát az egyik pillanatról a másikra megváltoztatni, az szükséges, hogy egy sejt sok, különböző idegrendszeri területtel álljon kapcsolatban. E felvetés a plazmid-mediálta GFP jelölés által megválaszolható volt. Felfedeztük, hogy a KR+KT sejtek szinte valamennyi, az érzelmi viselkedést meghatározó előagyi területre küldenek axonális kapcsolatokat. A prefrontális kéreg (személyiség, döntéshozás), a ventrális striátum (függőség és jutalmazási szabályozás), a bazális előagyi magok (figyelem szabályozás), a teljes amigdaláris magcsoport (félelem, szorongás, asszociatív tanulás), a hipotalamusz több magcsoportja, köztük az orexinerg sejtek (alvás-ébrenlét szabályozás), a szeptum, ventrális hippokampusz, entorhinális kéreg (eseményekhez kötött tanulási folyamatok), mind-mind a KT kalretinin sejtjeinek célterületei voltak . Sőt, spanyol együttműködő partnerünkkel, prof. Francisco Clasca segítségével azt is megvizsgáltuk, hogy egyetlen sejt rendelkezhet-e ennyire szerteágazó axonális hálózattal, vagy különböző KR+KT sejtcsoportot vetítenek egy-egy agyterületre. Egyedi megjelölt KR+KT sejtek idegvégződésit követve azt találtuk, hogy szinte valamennyi sejt nagyszámú (5-10) másik agyterületre küld axonokat (2. ábra), vagyis egyetlen sejt képes lehet szinkron aktivációs hullámot elindítani sok területen.
Rendelkeznek-e azonban a KR+KT sejtek a megfelelő szelektív bemenetekkel, amik révén tudomást szereznek az élőlény éberségi állapotáról? Vajon speciálisan ezek a sejtek kapják az agytörzs és a hipotalamusz ébredésért, motivációért felelős központjaitól a legtöbb bemenetet és nem a környező szomszédjaik? Következő lépésként, tehát két fontos aktiváló rendszert, a főleg agytörzsi eredetű glutamatáterg és az hipotalamikus eredetű orexinerg beidegzést megvizsgálva megállapítottuk, hogy ezen bemenetek szelektíven a KR+ sejteket idegzik be a talamuszban. A KR-t nem tartalmazó sejteket elkerülik. Ugyanezt a szelektív kapcsolatrendszer jelenlétét humán talamikus agyszövetben is igazoltuk, bizonyítva, hogy a KR+KT rendszer jelen van az emberi agyban is (3. ábra). Ez arra utal, hogy evolúciós szempontból ez egy nagyon ősi rendszer. Összegezve, kapcsolatrendszerük alapján így elmondható, hogy a KR+KT sejtek azért különlegesek, mert szelektív információkat kapnak az összes aktiváló felszálló idegpályától, a begyűjtött jeleket összegzik, majd ezeket az éberséggel-összefüggő információkat szétküldik az előagy legkülönbözőbb területeire. De hogyan vizsgálható ez a rendszer működése közben? Hogyan vizsgálhatjuk szelektíven a KR+KT sejtek működését? Hogyan léphetünk interakcióba e rendszerrel, tanulmányozzuk KR+KT sejtek serkentésének, gátlásának hatását az éberségre? Ezekhez a vizsgálatokhoz két dologra volt szükség. Egyrészt egy precízen számszerűsíthető éberségi szint változásra, másrészt a KR+KT idegsejtek szelektív aktiválására/gátlására. Az első problémára az alvás-ébrenléti átmenetet a legjobb megoldás. Ennek során ugyanis élettanilag és viselkedéstanilag is jól definiálható az éberségi szint változása. Az ébredés során az agyi elektromos aktivitás az alvásra jellemző lassú, ritmikus aktivitásból hirtelen kis amplitúdójú, rendezetlen aktivitásba vált. Ez az egér esetében, az emberhez hasonló módon, a koponyához rögzített EEG (elektroenkefalográfia) elektródákkal mérhető. Ezzel párhuzamosan az állat megmozdul, ami részint az izommozgás hirtelen megváltozásával (EMG, elektromiográfia), részint videofelvételek alapján detektálható. Ezek szerint pontosan tudunk jellemezni egy éberségi szint változást. De hogyan lehet a KR+KT sejtek aktivitását és az általuk keltett hatásokat vizsgálni? Erre a fent említett a sejttípus szelektív genetikai technikák mellett a XXI. század másik forradalmi, technikai vívmányára, a fény által aktiválható/gátolható idegsejtek módszerére, az optogenetikára (BOX 4) volt szükség.

Az optogenetika
Az optogenetika központi molekulája egy évtizedek óta ismert fehérje, a Chlamydomonas reinhardtii nevű ostoros moszat „channel-rhodopsin” nevű fehérjéje. E membránfehérje legfőbb tulajdonsága, hogy kék fény hatására megnyílik, és pozitív ionokat enged a sejt belsejébe, így depolarizálja azt. A moszat eredetileg az ostorának molekuláris motorját hajtja vele, így mozog a fény felé. Alig tíz éve azonban az agykutatók rájöttek, hogyha a moszattól kölcsönvett molekulát az idegsejtek membránjába építik és megvilágítják azt, az idegejt depolarizálódik és milliszekundumnyi precizitással akciós potenciált generál rövid fényimpulzusok hatására. Előállíthatók tehát a fény által hajtott idegsejtek. Ha ezt a nagyszerű technikát kombináljuk a fent említett genetikai technikákkal, máris adott egy idegsejtcsoport szelektív aktiválhatóságának lehetősége. Mi szükséges hozzá? Egy egértörzs, melynek adott idegsejt populációja CRE-rekombinázt fejez ki, egy plazmid, ami inaktív formában hordozza a channel-rhodopsint, egy kapilláris, amivel a megfelelő agyterületre juttatjuk a plazmidot. Így elérhetjük, hogy csak a CRE-rekombinázt kifejező idegsejtek membránjába jelenik meg a fény által aktiválható fehérje. Ha ezután egy hajszálvékony (0,1 mm) száloptikát beültetünk arra az agyterületre, ahol a channel-rhodopsint tartalmazó neuronok vannak, már csak egy pontosan szabályozható fényforrás (pl. lézer) kell és az idegsejtek aktivitását pontosan vezérelhetjük.
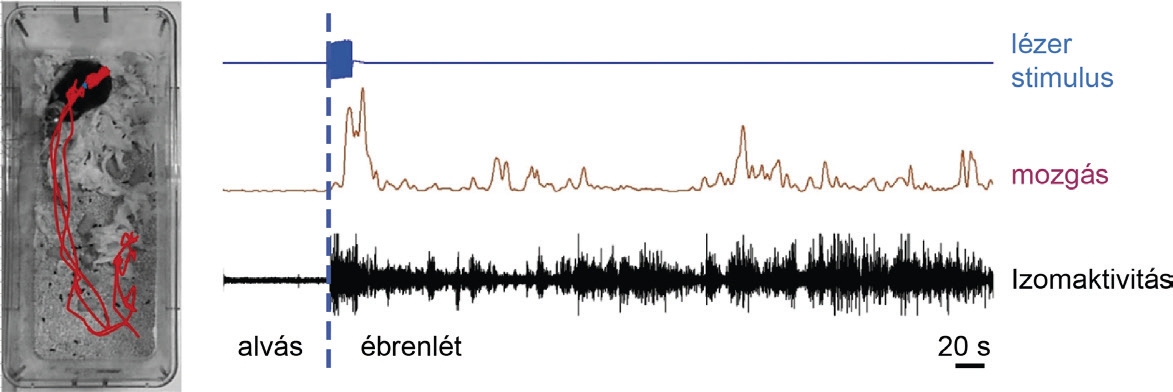
Az optogenetika eszköztára nagyon sokféle módon használható. Először is azonosítani tudjuk élő, szabadon mozgó állatban az adott idegsejt típust. Ha a sejtek kék fényre nagyon precízen és gyorsan aktiválódnak, akkor az adott populációhoz, esetünkbe a KR+ sejtekhez tartoznak. Szabadon alvó állatokban így megvizsgálhattuk, hogyan viselkednek a KR+KT sejtek felébredés során, illetve aktiválásuknak van-e ébresztő, azaz éberségi szint fokozó hatása. Az optogenetikailag azonosított 31 KR+KT sejt közül 29 növelte az aktivitását az felébredés során. Sőt, ezek közül 20 már a felébredés (izomaktivitás megjelenése) előtt 5-10 másodperccel elkezdte fokozni aktivitását (4. ábra), mely arra utalt, hogy a KR+KT sejtek aktiválódása és a felébredés között ok-okozati összefüggés lehet. Ennek közvetlen igazolására ugyancsak optogenetika segítségére támaszkodva a szabadon alvó egerek KR+KT sejtjeit aktiváltuk különböző intenzitású és hosszúságú fényimpulzussal. Legnagyobb megelégedésünkre a KR+KT hálózat szelektív aktiválásával valóban sikerült felébreszteni az alvó kísérleti állatokat (5. ábra). Érdekes módon az agyi EEG az aktiválás pillanatában megváltozott, ezt azonban csak néhány másodperccel később követte az izommozgás. Ez közvetlen igazolása volt annak, hogy a KR+KT sejtek először az előagyat ébresztik fel (EEG változás), a test felébredése ennek következménye (EMG aktivitás). Az ébredés mértéke attól függött, hogy milyen hosszú optogenetiai aktivációt alkalmaztunk. Ha csak rövidet (1 másodpercig, mp), akkor ún. mikroébredés következett be. Mikroébredés egy éjszaka során velünk is sokszor előfordul, hajnal felé 10 percenként akár 3-4-szer is. E rövid ébredési periódusok (2-30 mp) során átfordulunk a másik oldalunkra, de nem térünk öntudatra, tovább alszunk, és nem emlékszünk semmire az egészből. Az 1 mp-es KR+KT sejtaktiváció által kiváltott mikroébredések paraméterei teljesen megegyeztek a spontán mikroébredésekkel. Ez azt igazolja, hogy biológiailag releváns viselkedési választ váltott ki az általunk használt aktivációs fényimpulzus. Ha hosszabb ideig (10 mp) aktiváltuk a sejteket, az egér rendesen felébredt, éppúgy, ahogy magától tenné – és hosszú ideig ébren is maradt. Azt is megvizsgáltuk, milyen gyorsan érik el a KR+KT sejtek elágazó rostjainak impulzusai az előagy különböző részeit. A KR+KT sejtek optogenetikai stimulálása mellett párhuzamosan elvezettünk soksejt-aktivitást (multi-unit aktivitás, MUA) a KR+KT sejtek három legfőbb célterületéről. Kísérleteinket legújabb fejlesztésű, szilikon alapú sokcsatornás elektródák segítségével hajtottuk végre. Valamennyi agyterületen rövid késlekedési időt (néhány ezred mp) követően megjelentek a kiváltott MUA aktivitások, bizonyítva, hogy ez a rendszer képes gyors ingerület-továbbítására számos előagyi struktúra felé. Ezen eredmények tehát azt mutatták, hogy a KR+KT sejtek szabályozzák az ébredés folyamatát egy gyors, hatékony, elágazó pályarendszer segítségével. Utolsó kísérletünkben azt is megvizsgáltuk, hogy éber állapotban is van-e hatásuk az állatok éberségére. Ehhez egy olyan opszint használtunk fel, amely negatív ionok számára átjárható, így a fényimpulzusok hatására a sejtek aktivitása gátlódik. Egy nyílt téren azok az egerek, melyek KR+KT sejtjeit a gátló hatású opszin segítségével elcsendesítettük, csökkent motoros aktivitást, több rövid idejű megállást mutattak. Ezek a csökkent éberség jelei lehetnek, ami tovább erősíti KR+KT sejtekről kialakult képet.

Kereszttűzben a középvonali talamusz
Az elmúlt 3 év egyik legszélesebben kutatott agyterülete a KT. Jelentős felfedezések születtek igazolva, hogy a KT részt vesz a félelem, szociális interakció, drog-függőség vagy falánkság szabályozásában, nyitva hagyva a kérdést, hogy egy rendszer hogyan képes ennyi, látszólag nem összefüggő folyamatok irányítására. Úgy gondoljuk eredményeink választ adnak erre. A KR+KT hálózat összegezve az agytörzsi éberségi információkat, gyors axonális kapcsolataik révén üzen az előagynak, hogy „itt jön valami váratlan, új, érdemes figyelni”. Vagyis az agy éberségét fokozza, az agy állapotát változtatja meg. Annak megítélése, hogy milyen változásról van szó és milyen viselkedési válaszreakciót kell kialakítani, például ellenállhatatlan svédasztalról vagy valami rémületes helyzetről, már más agyterületek feladata. Eredményeink szerint a „fontosság” jelzése mellett ugyanez a rendszer felelős a legjelentősebb éberségi szint változás gyors kialakításáért, amikor az alvóállapotból éber állapotba megyünk át.
Gyökeresen más altatók
E rendszer felfedezése új megvilágításba helyezi azt a régi tapasztalatot, hogy az alvászavarok jelentős része a stressz miatt alakul ki. Hiszen most már rá lehet mutatni az agy egy jól körülhatárolható idegsejt hálózatára, melynek legfontosabb feladata, hogy reagáljon a külvilág állandó változásaira. A jelentős hatások, stresszes események így mély nyomot hagyhatnak bennünk ezen sejtek túlzott serkentődése révén. Elképzelhető tehát, hogy a folyamatos stressz ezen a hálózaton keresztül kihat az alvásunkra is. Így ha valamiképpen befolyásolni lehetne a hálózat működését, talán kezelhetővé válnának az alvászavarok, és a természetes alvás jellegzetességeit jobban visszaadó, újfajta altatókat fejleszthetnének ki. A Nature Neuroscience-ben most megjelent cikk két vezető szerzője már évek óta dolgozik együtt ezen a témán a Nemzeti Agykutatási Program (NAP) támogatásával. Mátyás Ferenc (aki éppen a NAP támogatásával alapította meg saját kutatócsoportját az MTA TTK-n) az egerek agyszövetének anatómiai vizsgálatát végezte, kísérletesen vizsgálta az e sejtekből kiinduló axonok elágazásait és szinkronaktivitását, és kollégájával, Kocsis Kingával mérte a hálózat aktivitását a szabadon alvó egér agyában. Acsády László nevéhez köthetők az optogenetikai kísérletek, melyben legfőbb segítsége Komlósi Gergely volt és az emberi szövetminták vizsgálata, amit Dávid Csaba végzett.
A cikk a Természet Világa 2019. januári (150. évf. 1. sz.) számában jelent meg.