A híres balti borostyánkövek az eocénben, mintegy 40-50 millió éve élt fák – főleg fenyőfák – gyantájából keletkeztek. Az úgynevezett borostyánerdők jellemzően száraz területeken álltak, ezért rejtélynek számított, hogy a borostyánkövekben megőrződött rovarok közel egynegyede miért vízirovar. E furcsa jelenség egyik lehetséges magyarázatát német és amerikai kutatók adták meg, amikor feltételezték, hogy a borostyánerdők tavak partján álló néhány (fenyő)fája viharok alkalmával a vízbe dőlhetett, s az így vízbe kerülő folyékony gyantába beleragadtak a vízirovarok. E hipotézist egy floridai mocsárerdőben ellenőrizték. Kiderült, hogy a vízbe folyó fenyőgyantába számos vízirovar tényleg beleragadhat, megteremtve ezzel a borostyánkővé fosszilizálódás egyik előfeltételét. E magyarázat szerint tehát „a hegy (fenyőfa) ment Mohamedhez (vízirovarhoz)”. Mi egy fordított logikájú, új magyarázatot javasoltunk, amely szerint „Mohamed ment a hegyhez”, vagyis a vizet kereső polarotaktikus vízirovar szállt rá a fenyőfatörzs sérüléseinél kifolyt friss gyantára, ami a vízszintesen poláros visszavert fényével vizet utánzott a polarizációérzékeny rovar számára. Cikkünkben ezt fejtjük ki részletesen.
Borostyánba zárva számtalan rovarfaj egyedére bukkantak már, amelyek közel egynegyede vízirovar [1]. Mivel a borostyán szárazföldi nyitvatermő fák megkövesedett gyantája, ezért rejtély volt, hogy miként került oly sok vízirovar a borostyánba. Az elsődleges vízirovarok (például vízibogarak – Coleoptera) lárvái és kifejlett formái is a vízben élnek, míg a másodlagos vízirovaroknak (például szitakötők – Odonata, lószúnyogok – Tipulidae, bögölyfélék – Tabanidae) csak a lárvái élnek vízben.
A vízirovarok borostyánba kerülését korábban többféleképpen magyarázták. Az első lehetőség, hogy a vízirovarok bármilyen okból is, de a rajzásuk és szétszóródásuk során kerültek a gyantába [2]. A második magyarázat szerint a gyanta belefolyt a fa vizet tartalmazó mélyedéseibe és az ott élő vízirovarok beleragadtak a gyantába [3]. Az a lehetőség is felmerült, hogy a friss gyantába a szél fújta bele a már elpusztult vízirovarok tetemeit [4]. Az is felmerült, hogy a gyanta a törzsön lefolyt a fa alatti vízbe, majd az ott élő vízirovarok beleragadtak a gyantacseppekbe [5].
Mindeddig csupán ezt az utolsó hipotézist támasztotta alá kísérletileg egy floridai mocsárerdőben végzett kutatás [5] . Cikkünk az első föltevésre épül, ami a vízirovarok aktív, szárazföld fölötti repüléséből indul ki, azonban korábban nem volt ismert, hogy konkrétan miért ragadtak bele a fatörzsekre kifolyt gyantába. Egyesek szerint a gyanta csillogó felszíne csalogathatta magához a vízkereső rovarokat [2] . Kutatási eredményeink e hipotézist támasztják alá azzal az újdonsággal, hogy a gyantás fatörzsek nem a csillogásukkal, hanem a visszavert fény vízszintes polarizációjával vonzották magukhoz a vízkereső, polarotaktikus vízirovarokat [6].
Ennek az az alapja, hogy a vízfelületekről általában vízszintesen poláros fény tükröződik és megannyi vízirovarfaj ennek érzékelésével ismeri föl a víztesteket [7]. A fény rezgéssíkját az összetett szemeik alsó tartományában található speciális, polarizációérzékeny fotoreceptorokkal érzékelik. Számos kísérletben kiderült, hogy több mint 350 vízirovarfaj sokkal erősebben vonzódik a vízszintesen poláros fényhez, mint a polarizálatlanhoz, valamint vízkeresésük az optikai környezet vízszintes polarizációján alapul és más tényezők (például szín, csillogás, szag, íz, hőmérséklet) másodlagosak [8].
Ha egy vízkereső, repülő vízirovar vízszintesen poláros fényt érzékel alulról, akkor leszáll az ilyen fényforrásra. Az elsődleges vízirovarok a víztest azonosítása után leszállnak a vízfelületre, vagy egyenesen fejest ugranak a vízbe. Ezzel szemben a másodlagos vízirovarok csak érintik a vízfelületet, aminek célja például az ivás, petézés vagy testük hőmérsékletének szabályozása. E reakciók során a rovarok legalább egyszer érintik a vizet, ami végzetessé válik, ha víz helyett mondjuk egy ragadós, gyantás fatörzsre szállnak.
Terepkísérletek
Terepkísérleteinket 2018 júniusában végeztük a Pilis hegységbeli Dömörkapunál csörgedező Bükkös-patak mentén, ami számos vízirovarfajnak ad otthont. Különböző színű olajtálcák és ragadós fatörzsek polarizációs tulajdonságait mértük és számszerűsítettük a rovarfogásaikat. Az 1. kísérletben egymástól 1 méterre négy műanyag tálcát (50 cm × 50 cm × 3 cm) helyeztünk egy aszfaltútra egyenes vonalban (1A ábra). A tálcák aljához 6 mm vastagságban fenyőkérget ragasztottunk, majd a tálcával együtt feketére, világos barnára, sötét barnára vagy fehérre festettük. A tálcákba 3 milliméter magasságig sárgás, átlátszó étolajat öntöttünk. E tálcák különböző színű kidőlt, ragacsos (gyantás) fatörzseket modelleztek. Az olajos felületekre szálló rovarok odaragadtak. A felületek sorrendjét félóránként véletlenszerűen változtattuk.
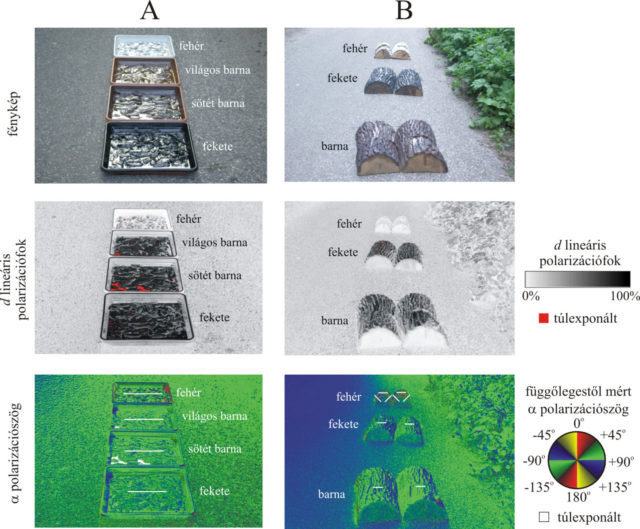
A 2. kísérletben barna, fekete és fehér félbevágott diófatuskókat helyeztünk az útra egymástól 1 m távolságra, egyenes mentén (1B ábra). Minden csoport (60 cm × 60 cm × 15 cm) az 1. kísérletben használt festékekkel lefestett négy féltönkből állt (30 cm × 30 cm × 15 cm). A diótuskók felületét színtelen, szagtalan, átlátszó, időjárásálló rovarragaccsal vontuk be.
A 3. kísérlethez három szomszédos függőleges törzsű juharfát választottunk ki közvetlenül a patak partján, amelyek közül az egyiknek fehérre, egy másiknak feketére festettük a törzsét a földtől mért 1 és 2 méteres magasság között (2. ábra). A festett felületeket és a festetlen harmadik barna fatörzs megfelelő területét a 2. kísérletben használt ragaccsal vontuk be.

A 4. kísérletben két kidőlt juharfa patakon közel vízszintesen átívelő barna törzsét ragasztóztuk be (3. ábra).
A ragadós tesztfelületek polarizációja
Az 1. kísérletben használt fekete, sötét barna, világos barna és fehér olajtálca vízszintesen poláros fényt vert vissza (1A ábra). A fekete és a két barna tálcáról visszaverődő fény d polarizációfoka 80 százaléknál nagyobb volt a függőlegestől mért 56,3°-os Brewster-szögből nézve. A fehér tálca csak gyengén poláros fényt tükrözött (d < 30 %). A fekete és barna tálcák erősen és vízszintesen poláros jele igen vonzó volt a vízkereső polarotaktikus vízirovaroknak, míg a fehér tálca nem volt vonzó.
A 2. kísérlet barna és fekete ragadós fatönkjeiről visszaverődő fény polarizációfoka ugyancsak nagyobbnak adódott 80 százaléknál, míg a fehér fatönkökről visszavert fényé kisebb volt, mint 15 százalék a Brewster-szögben (1B ábra). Mindemellett a visszaverődő fény rezgéssíkja nem volt mindig vízszintes, hanem különféle ferde irányok is előfordultak. A fekete és barna fatönkökről visszaverődő erősen és vízszintesen poláros fény vonzotta a vízkereső vízirovarokat, míg a fehér tuskók egyáltalán nem voltak vonzóak.
A 3. kísérlet függőleges ragadós fatörzsei ferdén, illetve függőlegesen poláros fényt vertek vissza (2. ábra). A legpolárosabb fényt a fekete (d < 50 %), a legkevésbé polárosat pedig a fehér (d < 15 %) fatörzs verte vissza, míg a barnáról visszaverődő fény polarizációfoka a kettő közé esett (d < 30 %) a Brewster-szögben. Mivel a visszavert fény rezgéssíkja mindhárom esetben jelentősen eltért a vízszintestől, e felületek nem voltak vonzóak a vízirovaroknak.

A 4. kísérlet két vízszintes kidőlt fatörzsének ragadós felülete nagy polarizációfokú (d > 60 %) fényt vert vissza a Brewster-szögben (3. ábra). Amikor e fatörzsek árnyékban voltak, azaz a diffúz égboltfény és a környező növényzetről eredő fény világította meg őket, a róluk visszaverődő fény rezgéssíkja vízszintes volt. Amikor viszont közvetlen napfény érte őket, a visszavert fény rezgéssíkja ferde volt. Az előbbi esetben a vízirovarok számára vonzóak voltak, ám az utóbbiban nem.
Csapdázott rovarok
Az 1. kísérletben a fekete, sötét barna, világos barna és fehér olajtálcák rendre 989, 461, 273 és 5 árvaszúnyogot fogtak, amely számok szignifikánsan különböznek egymástól.
Tehát minél sötétebb volt egy tálca, annál több polarotaktikus árvaszúnyogot csalogatott magához. A 2. kísérletben csupán 5 és 11 kérészt fogtak a sötét barna és fekete ragadós fatuskók.
Az 1. táblázat a 3. és 4. kísérletben a ragadós fatörzseken csapdázódott vízirovartaxonokat tartalmazza. A barna, fekete és fehér függőleges fatörzsek rendre 10, 9 és 9 különböző taxont fogtak. A három vízszintes fatörzsön összesen 13, míg a vízszintes fatörzseken 14 féle taxon csapdázódott (4. és 5. ábra). A függőleges fekete és barna fatörzs gyakorlatilag ugyanannyi egyedet fogott (Nbarna = 26, Nfekete = 23), míg a függőleges fehér szignifikánsan kevesebbet, nagyjából feleannyit (Nfehér = 11). A vízszintes ragadós barna fatörzsek összesen Nvízszintes = 60 rovart csapdáztak (4. és 5. ábra), és a felületi rovarsűrűségük közelítőleg megegyezett a függőleges barna fatörzsekével.
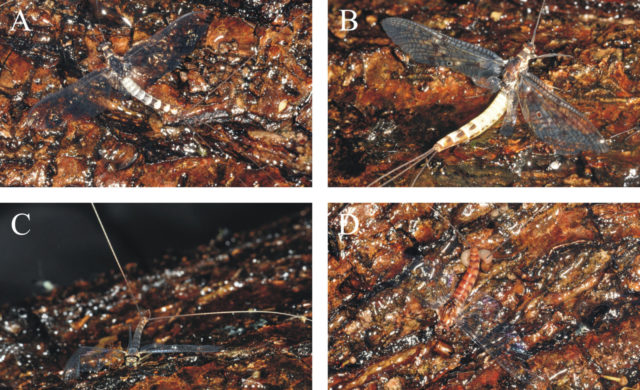
A függőleges fatörzseken egyetlen imágó kivételével mind a 14 csapdázódott kérész szubimágó volt. A szubimágó állapot a hímek és nőstények fejlődésének utolsó előtti szakasza, amiből egy végső vedlés vezet a párzóképes imágó állapotba. Így tehát a függőleges fatörzseken párzásra képtelen kérészek csapdázódtak. A vízszintes fatörzsek nőstény és hím imágókat fogtak és a nőstények mellett gyakran petecsomókat is megfigyeltünk.
Megtévesztett kérészek
Mivel a borostyánkövekben gyakoriak az olyan rovarfajok imágói, amelyeknek legalább a lárvája vízben fejlődik, ezért jogosan tételezhetjük fel, hogy számos tó és patak tarkította a korabeli borostyánerdők vidékét. A mai fenyőfákból származó gyanták közel 50 százaléka megszilárdul, miután kifolyt a fakéregre [9]. Így a vízirovarok borostyánban való jelenléte jól magyarázható azzal, hogy a fából kifolyt, még folyékony gyantába ragadtak bele a repülő rovarok. Egy elképzelés szerint a repülő vízirovarokat a gyanta csillogása csalogathatta magához [2]. De nyitott maradt a kérdés, hogy vajon miért lehetett optikailag olyan vonzó a gyanta a vízirovarok számára, továbbá kísérleti bizonyíték sem született ezen elképzelés igazolására. Kísérleteinkkel azt tanulmányoztuk, hogy a repülő vízirovarok számára a gyantás fatörzsek polarizációja milyen szerepet kap e vonzásban.
Eredményeink azt mutatták, hogy kizárólag a vízszintes ragadós fatörzsek képesek vonzani a vízirovarokat a róluk visszaverődő fény vízszintes polarizációjával. Függőlegesen vagy ferdén poláros fényt visszaverő, nem vízszintes fatörzsek nem vonzzák őket, így azokon nem is ragadnak bele a gyantába. Annak oka, hogy a különböző színű fatörzsekről eltérő polarizációfokú fény verődik vissza, a fényerősségben keresendő: minél sötétebb egy felület egy adott színtartományban, annál nagyobb a róla visszaverődő fény polarizációfoka.
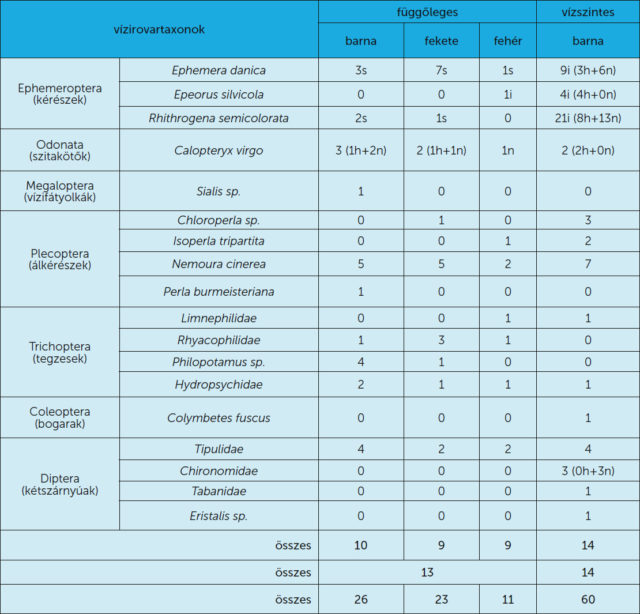
Kísérleteink szerint a csapdázódott rovarfajok jól tükrözik a helyi rovarfauna összetételét. A függőleges fatörzsekre csak kevés rovar ragadt, továbbá a felület világos színe jelentősen lerontotta a vonzóképességet. A függőleges törzsek nem váltottak ki olyan erős vonzást, ami a rovarok tömeges csapdázását eredményezhette volna. Habár a vízszintes és függőleges fatörzsek egyaránt fogtak a legtöbb helyi rovarfajból, azonban a legérdekesebb jelenséget a kérészeknél figyeltük meg: a függőleges fatörzseken csak szubimágók csapdázódtak, míg a vízszinteseken kizárólag imágók (1. táblázat). Mindennek az az oka, hogy a vízszintes felületekről vízszintesen poláros fény verődik vissza, ami képes megtéveszteni a polarotaktikus vízirovarokat, így a kérészeket is [10]. A függőleges fatörzsekről nem vízszintes rezgéssíkú fény verődött vissza, ezért ezek nem vonzották a vízirovarokat. Kikelésük után a kérész szubimágók általában olyan tereptárgyat keresnek a víz közelében, ahol párzóképes imágóvá tudnak vedleni. Ezért a függőleges fatörzseken való csapdázódásuk feltehetően a véletlen műve volt. Hasonló véletlen csapdázódás áldozata lehetett az a kisasszony-szitakötő (Calopteryx virgo) is, amit éppen átváltozás közben ért a vég a ragadós fatörzsön (5A ábra) olyan pozitúrában, amelyben valódi borostyánban is előkerült már szitakötő [11].

Terepkísérleteinkkel tehát megállapítottuk, hogy a vízszintesen poláros fényt visszaverő ragadós fatörzsek vonzzák a vízkereső repülő vízirovarokat, továbbá sokféle (vízi és szárazföldi) rovar könnyen ráragadhat egy vízfolyás melletti ragadós fatörzsre. E megállapítások elégséges föltételei azon hipotézisünknek, miszerint a polarotaktikus vízirovaroknak vonzóak a gyantás fatörzsek, amennyiben vízszintesen poláros fényt vernek vissza.
Manapság rengeteg olyan mesterséges felület létezik, ami a vízirovarok szemében víznek minősül, mivel erősen és vízszintesen poláros fényt vernek vissza. Ilyenek például az olajtározók, aszfaltutak, földre terített fekete műanyag fóliák, üvegfelületek, sötét autókarosszériák, fekete márványkövek és napelemtáblák. Általában vonzást vált ki a polarotaktikus vízirovarokból minden sötét és fényes felület, ami erősen és vízszintesen poláros fényt ver vissza. E vizuális vonzásnak sokszor végzetes a hatása az érintett rovarokra, mivel a lerakott petéik kiszáradva elpusztulnak a száraz felületeken. E jelenséget poláros fényszennyezésnek nevezzük [12]. Így tehát a vízirovarok fagyantában történő csapdázódására adott magyarázatunk a poláros fényszennyezés egy ősi formája, habár hatása oly csekély lehetett, hogy nem okozhatott számottevő hátrányt a vízirovar-populációknak.
HORVÁTH GÁBOR – EGRI ÁDÁM – BENNO MEYER-ROCHOW – KRISKA GYÖRGY
IRODALOM
[1] Weitschat W., Wichard W. (2010) Baltic amber. In: Biodiversity of Fossils in Amber from the Major World Deposits. Penney D. (ed.) Siri Scientific Press: Manchester, pp. 80-115
[2] Poinar G. O. (1992) Life in Amber. Stanford University Press
[3] Miller K. B. (2003) The unusual occurrence of aquatic beetles in amber, Copelatus aphroditae Balke, n. sp., and C. predaveterus Miller, n. sp. (Coleoptera: Dytiscidae: Copelatinae). Proc. Entomological Soc. Washington 105: 809-815
[4] Keyser D., Weitschat W. (2005) First record of ostracods (Crustacea) in Baltic amber. Hydrobiologia 538: 107-114
[5] Schmidt A. R., Dilcher D. L. (2007) Aquatic organisms as amber inclusions and examples from a modern swamp forest. PNAS 104: 16581-16585
[6] Horváth G., Egri Á., Meyer-Rochow V. B., Kriska G. (2020) How did amber get its aquatic insects? Water-seeking polarotactic insects trapped by tree resin. Historical Biology doi: 10.1080/08912963.2019.1663843
[7] Schwind R. (1991) Polarization vision in water insects and insects living on a moist substrate. J. Comparative Physiol. A 169: 531-540
[8] Horváth G., Csabai Z. (2014) Polarization vision of aquatic insects. In: Horváth G. (ed.) Polarized Light and Polarization Vision in Animal Sciences. Springer: Heidelberg, pp. 113-145
[9] Henwood A. (1993) Recent plant resins and the taphonomy of organisms in amber: a review. Modern Geology 19: 35-59
[10] Egri Á., Pereszlényi Á., Farkas A., Horváth G., Penksza K., Kriska G. (2017) How can asphalt roads extend the range of in situ polarized light pollution? A complex ecological trap of Ephemera danica and a possible remedy. J. Insect Behavior 30: 374-384
[11] Wichard W., Gröhn C., Seredszus F. (2009) Aquatic Insects in Baltic Amber. Remagen Verlag: Kessel
[12] Horváth G., Kriska G., Malik P., Robertson B. (2009) Polarized light pollution: a new kind of ecological photopollution. Frontiers in Ecology and the Environment 7: 317-325
A cikk a Természet Világa 2020. júniusi számában (151. évf. 6. sz.) jelent meg.