A környezeti tényezők, köztük a fény, a páratartalom vagy éppen a hőmérséklet döntően befolyásolják a növények mindennapi életét. Köztük is a fény az egyik olyan kulcsfontosságú tényező, mely nemcsak a növények optimális növekedését és fejlődését szabályozza, hanem fontos szerepet játszik a növények stresszválaszaiban, köztük a növényi kórokozókkal vagy a káros környezeti hatásokkal szembeni védekezésben is.
A fény kvantitatív és kvalitatív tulajdonsággal is bír, melyeket a növények speciális fotoreceptorok (például vörös fényt érzékelő fitokrómok vagy a kék fényt érzékelő kriptokrómok) segítségével érzékelnek. A fotoreceptorokkal történő fényérzékelés nemcsak a növények növekedését és fejlődését, hanem védekezésüket is döntően befolyásolja. Emellett a fény biztosítja a szükséges energiát a növények fotoszintéziséhez is, mely során az energiáját felhasználva szerves anyagokat (szénhidrátokat) állítanak elő. Éppen ezért a megfelelő fényviszonyhoz képest a magas- és alacsony fényintenzitás, a fény összetételének változása, az árnyék vagy a sötétség, vagy akár a fényperiódus hosszának és időszakának megváltozása eltérő növényi stresszválaszokat okozhatnak.
Ezek alapján kérdésként merülhet fel, vajon mi történik a fény hiányában (például éjszaka vagy tartós sötétség alkalmával) a növényekben? Milyen élettani, biokémiai és molekuláris biológiai változások zajlanak a fényhez képest? Mindez hogyan befolyásolja a növények védekezését a változó környezeti kihívásokkal szemben?
Janus-arcú molekulák
A római mitológiából ismert Janus a kezdet és a vég istene volt, akit két arccal ábrázoltak utalva az általa képviselt kettősségre. Az élőlényekben, így a növényekben is vannak olyan kettős természetű vegyületek, melyek egyszerre játszanak szerepet „békében és háborúban”. Ezek közé tartoznak egyes reaktív oxigénformák (ROF), melyek párosítatlan elektronnal rendelkeznek és ezáltal képesek gyorsan elektront elvonni környezetükből. Közéjük tartozik például a szuperoxid-gyök anion (O2•–) és a hidrogén-peroxid (H2O2) is. A szuperoxid-gyökanion rendkívül reakcióképes, féléletideje 1-4 μs és migrációs távolsága 30 nm a sejten belül. Ezzel szemben a hidrogén-peroxid féléletideje több, mint 1 ms és migrációs távolsága akár az 1 μm-t is meghaladhatja. A hidrogén-peroxid természetére igaz talán leginkább a kettősség. Sokáig ugyanis káros molekulának tartották, mára azonban bebizonyosodott, hogy nagy koncentrációban valóban károsítja a sejteket alkotó fehérjék, lipidek és nukleinsavak szerkezetét és ezáltal működésüket, valamint bebizonyították, hogy szerepet játszik a programozott sejthalál kiváltásában, mely segítségével a növény meg tudja fékezni egyes kórokozók továbbterjedését a fertőzés helye körüli sejtek pusztulásával a levelekben. Ugyanakkor felfedezték azt is, hogy kis koncentrációban nagyon fontos szerepet játszik a sejtek jelátviteli folyamataiban. Kimutatták ugyanis, hogy a hidrogén-peroxid többek között döntő szerepet játszik a sejtciklus, a növekedés és fejlődés, az öregedés vagy éppen a kórokozók behatolását gátló sztómazárás szabályzásában.

Reaktív oxigénformák számos folyamat során keletkezhetnek a növényi sejtek több részén (például mitokondrium, kloroplasztisz, peroxiszóma, apoplaszt). Keletkezésüket mind a környezeti stresszorok (például nehézfémstressz, szárazság, sóstressz, hőmérsékleti stressz, UV-B sugárzás), mind pedig a kórokozók (baktériumok, gombák, vírusok) általában fokozzák, így a fény a kloroplasztiszokban található elektrontranszportlánc túlműködése során jelentősen befolyásolhatja azt. Éppen ezért a növények a ROF semlegesítése céljából nagy mennyiségben szintetizálnak különböző enzimatikus és nem-enzimatikus antioxidánsokat.
Antioxidánsok akcióban
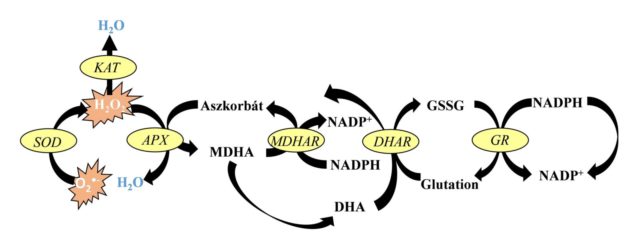
A ROF semlegesítésében kitüntetett szerepet játszanak a Foyer-Halliwell-Asada ciklus enzimatikus és nem-enzimatikus komponensei. Az enzimatikus antioxidánsok közül kiemelkedő a szuperoxid-dizmutáz (SOD), a kataláz (KAT) és az aszkorbát-peroxidáz (APX) szerepe. A SOD-ok olyan fémtartalmú enzimek, melyek a szuperoxid-gyökanion átalakítását végzik a kevésbé toxikus hidrogén-peroxiddá és oxigénné. A SOD-enzimeket fém-kofaktoraik alapján csoportosíthatjuk: mangán (Mn), vas (Fe) és réz/cink (Cu/Zn) SOD-enzimekre. A Mn-SOD a mitokondriumokban és a peroxiszómákban, az Fe-SOD a kloroplasztiszokban és a peroxiszómákban, a Cu/Zn-SOD pedig a kloroplasztiszokban, a peroxiszómákban, a citoszólban és az apoplasztban fordul elő. A keletkezett hidrogén-peroxid elbontását a legnagyobb mértékben a KAT és az APX végzi. A hemcsoportot tartalmazó, tetramer szerkezetű KAT elsősorban a peroxiszómákban található meg. Az APX azonban a kloroplasztiszokban, a citoszólban és egyes mikrotestekben is előfordul. A két hidrogén-peroxidot semlegesítő enzim között további különbség, hogy míg a KAT kis affinitással kötődik a hidrogén-peroxidhoz, amit nagy mennyiségben képes semlegesíteni, addig az APX nagyobb affinitással rendelkezik és az alacsonyabb H2O2-szinteket szabályozza működésével a növényekben.
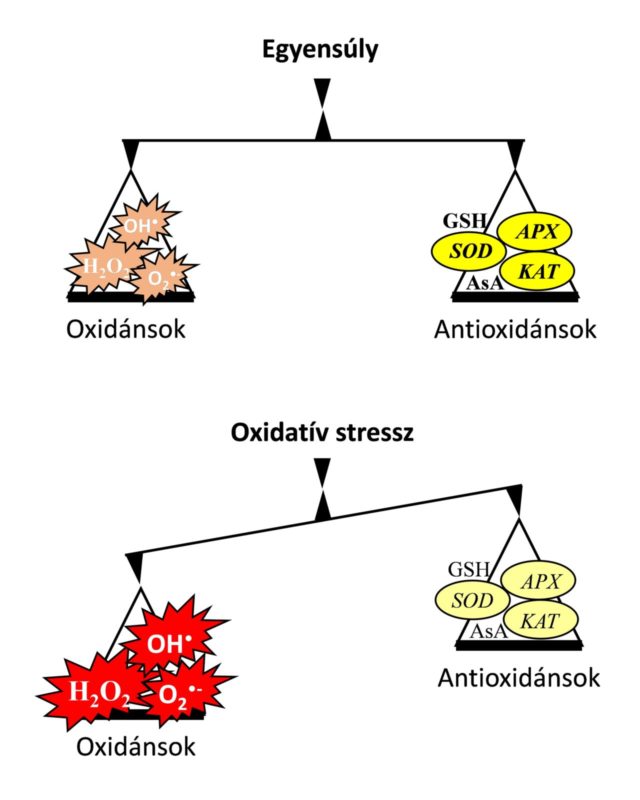
A nem-enzimatikus antioxidánsok közül kulcsszerepe van az aszkorbinsavnak (AsA) és a glutationnak (GSH). Az aszkorbinsav mennyisége a kifejlett fotoszintetizáló levelekben és egyes gyümölcsökben a legmagasabb a többi növényi szervhez képest. Az aszkorbinsav 30-40 százaléka ugyanis a kloroplasztiszokban található, de szintézisében és oxidált formájának regenerálásában a mitokondriumok vesznek részt. Az aszkorbinsav a kloroplasztiszok mellett megtalálható még a sejtmagban, peroxiszómákban, a citoszólban, a vakuólumban és az endoplazmatikus retikulumban is. Az aszkorbinsav képes közvetlenül reakcióba lépni a szuperoxid-gyök anionnal, valamint az APX működése révén a hidrogén-peroxidot vízzé alakítani. Az aszkorbinsav mellett a glutation, mely egy tripeptid (γGlu-Cys-Gly), egy másik jelentős antioxidáns, mely kénraktárként is funkcionál a növényekben. A redukált glutation szinte minden növényi sejtkompartmentumban megtalálható, köztük a citoplazmában, endoplazmatikus retikulumban, vakuólumban, mitokondriumban, kloroplasztiszban, peroxiszómában és az apoplasztban is. A glutation szintén fontos szerepet játszik a különböző ROF-molekulák, köztük a hidrogén-peroxid semlegesítésében. Szerepét leírták számos biológiai folyamatban, többek között a növekedés és fejlődés szabályozásában, a sejtdifferenciálódás során, a sejthalál és az öregedés szabályozásában, valamint különböző kórokozókkal szembeni ellenállás és a környezeti stresszorokkal szembeni tolerancia kialakítása során.
Noha az antioxidánsok szerepe a növények életében megkérdőjelezhetetlen, mégis egyelőre kevés tudományos adat áll rendelkezésre a különböző stressztényezők hatására bekövetkező változásaikról eltérő fényviszonyok mellett. A növényi antioxidánsrendszer enzimatikus és nem-enzimatikus komponenseinek napszak- (cirkadián óra) valamint fényfüggő szabályozásának ismerete még nem teljesen feltárt.
A titokzatos éjszaka
A fény és a sötétség folyamatos változása által az élőlények belső cirkadián órája számos molekuláris, fiziológiai és metabolikus változást szabályoz. A növényekben a fényperiódus alatt felhalmozott szénhidrátok felhasználása szintén a cirkadián óra szabályzása alatt áll. A napi ritmus során a felkelő Nap hajnali kék fénye stimulálja a levelekben található gázcserenyílások nyitódását, míg a kék fény a vörös hullámhossz-tartományba eső fénysugarakkal együtt a szén-dioxid asszimilációját serkenti a fényperiódusban. Este, sötétben a gázcserenyílások ismét zárva vannak, mely egyben az elsődleges védelmi vonalat is jelenti a különböző növényi kórokozókkal szemben. Éppen ezért a kórokozók támadásának időpontja és a fény jelenléte vagy éppen hiánya kulcsfontosságú a növények védekezésében. Ugyanakkor jól ismert, hogy számos baktérium vagy gomba fertőző hatását a sötétség segíti, míg a fény közvetlenül gátolhatja, mint például a gombaspórák csírázását, a fonalak növekedését és a gombafertőzés hatékonyságát. Ismert, hogy a kórokozók a fertőzés helyén ROF termelődését váltják ki a növényi sejtekben, mely hozzájárul a helyi és szisztemikus védekezés indukciójához vagy éppen a programozott sejthalál kiváltásához. Kevésbé kutatott azonban, hogy a ROF képződésében és semlegesítésében szerepet játszó antioxidánsok szintjei és aktivitása hogyan változhat a különböző napszakokban és a sötétben.
Érdekes, hogy pár tudományos mérés igazolta, hogy kontroll körülmények mellett az antioxidáns enzimek és nem-enzimatikus antioxidánsok aktivitása és szintje eltér a különböző napszakokban, fényben és sötétben. Azaz az antioxidánsok mennyiségét és aktivitását nemcsak a növényfajták, a szervek és szövetek vagy a különböző fejlődési stádiumok és stressztényezők befolyásolják, hanem a napszak, így a fény és a sötétség is. Paradicsomnövények levelében megfigyelték például, hogy a SOD aktivitása a fényszakaszban a napfelkeltétől folyamatosan nő egészen a sötétszakasz kezdetéig, majd drasztikusan lecsökken a hajnali órákban. Hasonló változást mértek a KAT-enzimaktivitás esetében is. A két napszak között több, mint másfélszeres a különbség a két vizsgált antioxidáns enzimaktivitásában. Mások azt is kimutatták, hogy a Fe-SOD és Mn-SOD enzimeket kódoló gének kifejeződése este a sötétszakaszban jelentősen lecsökken. A SOD-enzimhez képest a KAT-izoenzimeket kódoló gének átíródása változatosabb képet mutat a különböző növényfajokban. Míg egyes gének kifejeződése a nap fényszakaszában folyamatosan nő és este a sötétben csökken (például kukorica CAT3, dohány CAT2 és CAT3), addig más KAT-gének átíródásának maximuma a reggeli órákban (például paprika CAT1, lúdfű CAT2) vagy koraeste (például lúdfű CAT3) figyelhető meg. Az APX-enzim aktivitása és a kódoló gének kifejeződése szintén erősen fényfüggő. Kimutatták dohányban, hogy a citoszólikus cAPX1 gén átíródása a nap fényperiódusában nő, míg este a sötétben ez jelentősen alábbhagy. Hasonló tendenciát figyeltek meg a kloroplasztikus sAPX és tAPX gének esetében is.
A nem-enzimatikus antioxidánsok esetében az enzimatikus antioxidánsokhoz hasonló változásokat figyelhetünk meg. Az aszkorbinsav magas reggeli szintjét mérték bükkfa levélben, ami a nap fényperiódusa alatt folyamatosan emelkedett, és amely délután mutatott maximumot, amikor a legmagasabb volt a napsugárzás intenzitása. Ezt a tendenciát figyelték meg alma levelében és termésében, valamint paradicsomban is. A glutation kapcsán végzett megfigyelések alapján a glutationtartalom magasabb volt napközben a fényen, mint éjszaka például luc- és kanári fenyők tűleveleiben. A fény pozitív szerepét igazolták borsónövény levelének sejtjeivel, ahol 10 perc hosszúságú sötétség hatására nem, viszont 10 perc hosszúságú fény alkalmazása következtében jelentősen megnövekedett a sejtek aszkorbinsav- és glutationszintje, a növekvő ROF-produkció mellett.

Ezek a mérések alátámasztják, hogy az enzimatikus és nem-enzimatikus antioxidánsok aktivitása és szintjei nem állandók a különböző növények leveleiben és terméseiben, hanem általában a fény hatására nappal emelkednek, míg sötétben éjszaka csökkennek, mely azonban jelentősen befolyásolhatja a növények védekezését egyes kórokozókkal szemben.
Összefoglalva megállapítható, hogy mind a napszak (nappal/éjszaka), mind pedig a fény jelenléte vagy hiánya döntő jelentőségű a növényi stresszválaszok finomhangolásában. Éppen ezért a ROF keletkezésének és lebontásának szabályozása, valamint a különböző enzimatikus és nem-enzimatikus antioxidánsok védekezésben szerepet játszó hatásainak vizsgálata és megértése nemcsak a gyakorta tanulmányozott fényszakaszban, hanem a sötétszakaszban, az éjszakai periódusban vagy mesterségesen előidézett sötétítés során is különösen fontos lehet, hogy pontosabban megismerhessük a növények mindennapi életét és a kórokozókkal folytatott küzdelmeit. E folyamatok felderítése és pontosabb megértése új perspektívákat kínálhat a növénybiológiai kutatások és az agrárium számára egyaránt.
POÓR PÉTER
Nyitóképünk: Az éjszaka számos kihívást tartogat a növények számára. Számos növényi kórokozó és növényevő aktívabb este, mint nappal.
A munka a Nemzeti Kutatási, Fejlesztési és Innovációs Hivatal (NKFIH FK 124871) és az Innovációs és Technológiai Minisztérium ÚNKP-19-4-SZTE-86 kódszámú Új Nemzeti Kiválóság Programjának szakmai támogatásával készült. Poór Pétert a Magyar Tudományos Akadémia Bolyai János Kutatási Ösztöndíja támogatja.
A cikk a Természet Világa 2021. februári számában (152. évf. 2. sz.) jelent meg.