|
Sejtmozgás és mintázatképződés Beszélgetés Czirók Andrással Az élő szervezetek tele vannak
mintázatokkal. Gondoljunk a gerinc csigolyáira, a tüdő légjárataira, az
ujjainkra vagy az élőlények külső jegyeire, a zebra csíkjaira és a pillangók
szárnyaira. "De nem is kell, hogy többsejtű élőlényeket vizsgáljunk, már
az egysejtű mikroorganizmusok is nagyon érdekes önszerveződő mintázatokat
tudnak képezni" - mondja Czirók András, aki másodéves egyetemista kora
óta figyeli őket. Ugyancsak jellegzetes mintázatok alakulnak ki szövetek
képződésekor - ilyenkor egy embrió testi sejtjei vándorolnak, hogy a kialakuló
szervezetben jól definiált helyüket megtalálják. Czirók András, az ELTE
fiatal kutatója a sejtek követésére szolgáló videomikroszkópos technika
kidolgozásáért és alkalmazásáért 2006 decemberében Nemzetközi Gábor Dénes-díjat
kapott.
- Mennyiben tér el a biológiai mintázatok leírása a szervetlen mintázatokétól? - Sejteket, szöveteket vagy még nagyobb rendszereket vizsgáltok? - Három téma köré csoportosíthatók a kutatásaink. Az első, a mikroorganizmusok telepképzése és mintázatképzése visszanyúlik a doktori iskolában végzett munkámhoz, amelyen Vicsek Tamás professzorral dolgoztunk. Ott az volt a kérdés, hogyan hat a külső, fizikai környezet ezekre az egymáshoz nagyon hasonló és egymással is kölcsönható élőlényekre. Például, ha a tápanyag egy gélen keresztül diffundál hozzájuk, akkor nagyon érdekes, fraktálszerű alakzatok alakulnak ki. Ilyenkor az egyes sejtek viselkedését alapvetően meghatározza egy fizikai diffúziós folyamat. Előfordulhat az is, hogy egy folyadék felszínén, a nagy oxigénkoncentráció miatt összegyűlnek a mikroorganizmusok. A folyadék sűrűsége ezen a helyen megnő, így hidrodinamikai instabilitások alakulnak ki. Ennek következtében az egész víztömeg áramolhat, ez a biokonvekció. Az is nagyon érdekes, hogy ha ezek a mikroorganizmusok elég nagy sűrűségben vannak jelen, akkor valahogy összehangolják a mozgásukat. Mindegyikük szeretne mozogni, de nem tudnak átmenni egymáson, így különleges áramlási mintázatok alakulhatnak ki, amelyek különböznek a jól ismert, folyadékok hidrodinamikája által leírt áramlási típusoktól. Ennek a munkának a szerves folytatása lett a másik két téma, a sejtmozgás és az embriogenezis vizsgálata. Sejtmozgással azért kezdtünk el foglalkozni, mert a szöveti mintázatképződésnek az az egyik hajtóereje, hogy a sejtek mozogni tudnak. Másrészt ez egy nagyon érdekes biofizikai probléma, hiszen a sejt fizikai test, amelyben a sejtváznak, a citoszolnak és a sejtmembránnak eltérő mechanikai tulajdonságai vannak. Ahhoz, hogy a sejt haladni tudjon, ezeket meg kell változtatni, erőket kell rájuk kifejteni. A sejtmozgás tehát olyan probléma, ahol a biofizika szorosan kapcsolódik a biokémiához, a sejtbiológiához és maga a sejtmozgás az alapja sok más mintázatképző rendszernek is.
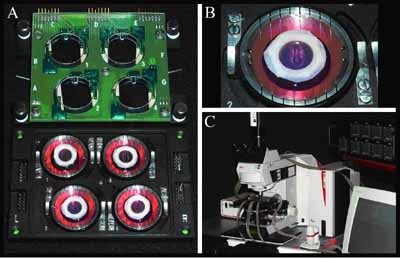
Inkubátor és videomikroszkóp madárembriók megfigyelésére. A tojásból kivett embriók egy tápoldat felszínén fejlődnek optimális optikai körülmények között A harmadik témám, az embriófejlődés során kialakuló anatómia ugyancsak mintázatképzési probléma. A képződő szerkezet sokkal jobban meghatározott, mint a mikroorganizmusok telepei, elsősorban a sokrétű, génexpressziós mintázatokat is magukban foglaló visszacsatolások miatt. Speciális gének expressziója definiálja az embriók különböző részeit és ez egyben behatárolja a sejtek „viselkedését”. Azt gondoljuk azonban, hogy ezen felül léteznek ún. emergens mintázatok is, amiket nem közvetlenül génexpressziós mintázatok határoznak meg. Erre lehet jó példa a meleg vérű gerincesekben megjelenő korai érhálózat: ezt körülbelül ezer endotél sejt alakítja ki az embriófejlődés első szakaszaiban. A sejtek kezdetben véletlenszerűen helyezkednek el, utána aggregálnak, nyúlványokat növesztenek, majd hálózatot képeznek. Ebből az elsődleges hálózatból később bizonyos helyeken főerek alakulnak ki, és ezekben már növekedési faktorokon keresztül génexpressziós mintázatok határozzák meg az alkotó sejtek számát. Mindenesetre úgy tűnik, hogy a különféle embriókban megjelenő változatos, elsődleges mintázat önszerveződő módon jön létre. - Mit jelent az "önszerveződés" egy biológiai rendszerben? - Az önszerveződés fogalmát a fizikusok előszeretettel használják. Nagyjából azt értjük alatta, hogy sok, nagyon hasonló alkotóelem kölcsönhatása egy nagyobb skálán olyan elrendeződést kelt, amely lényegesen különbözik attól, amit az alkotóelemek magukban, kölcsönhatás nélkül hoznának létre. Például, ha veszünk száz sejtet és jó messzire helyezzük őket egymástól, akkor önmagukban csak bolyonganak. Ha nagy számban vannak jelen, akkor össze tudnak tapadni, sőt bizonyos sejtek nem kupacot, hanem hálózatot hoznak létre. Ehhez járulnak a különféle biológiai visszacsatolások, hiszen a kialakult struktúrát az embriónak vagy a fejlődő szövetnek kontrollálnia kell. A túl kevés sejtet tartalmazó részekre újabb sejtek vándorolnak vagy az ott lévő sejtek osztódnak, a túl sűrű helyekről pedig a sejtek elvándorolnak. Ezeknek a folyamatoknak a részletes mechanizmusát még nem nagyon értjük. Azt tudjuk, hogy felnőtt állatban az oxigéngradiens "kemoattraktánsként" tud viselkedni érnyúlványok benövéséhez. Ezenkívül vannak olyan növekedési faktorok, amelyeknek a gradiense szintén vonzhatja ezeket a csúcsokat. Például bizonyos tumorok termelnek egy VEGF nevű molekulát (vascular endothelial growth factor, vaszkuláris endotél növekedési faktor), ami a szövetben diffundálni képes, és az így kialakuló gradiensek vonzó irányokat jelölnek ki a sejtek számára. Embriókban viszont még nem tudjuk, hogy az erek kezdeti mintázatát pontosan milyen mechanizmusok hozzák létre. Feltehetően az embriókban is vannak rövid skálájú gradiensek, amelyek orientálják a sejtek elrendeződését, de kísérletileg egyelőre nem sikerült bizonyítani, hogy ebben az esetben is a VEGF gradienséről lenne szó. Annyit lehet tudni, hogy ha megnézzük a szövetet, például tömegspektrometriával, akkor a VEGF eléggé egyenletesen oszlik el. Ugyanakkor lehetséges, hogy az extracelluláris mátrix csak bizonyos helyeken teszi a sejtek számára elérhetővé ezt a növekedési faktort. Mi egy „minimalista” megközelítési módot használunk, tehát azt mondjuk, hogy elég sok olyan fizikai effektus lehetséges, aminek eredményeként egy hálózat önszerveződésre képes, nem feltétlenül szükséges a növekedési faktorokhoz fordulnunk. Azt találtuk például, hogy ha egy hálózat bizonyos részén kevés a sejt, akkor azok a sejtek nagyon elnyúltak. Az elnyúlt sejtek sejtvázáról feltételezhetjük, hogy merevebbek, mert mechanikai feszültség alatt vannak. Számos sejttípusról ki lehet mutatni, hogy képesek érzékelni a környezet mechanikai tulajdonságait. Tehát például egy mechanikailag megfeszített sejt felszíne kedvező tapadási felszín lehet a környező sejtek számára. - Néhány sejt véletlenszerűen megnyúlik, és arra épülnek a többiek? - Igen, így képzeljük el egy sejtcsoportokat összekötő, többsejtű "híd" létrejöttét és későbbi stabilizálását is. Azon a helyen, ahol kevés a sejt, a sejtek megnyúlnak, és az így létrejövő vonzó felszínre érkeznek a többiek. Ezek persze elképzelések, amelyek a rendszer megértésére szolgának. Kísérleteket, majd ezek alapján modellszámításokat végzünk. A modell ahhoz szükséges, hogy kiderítsük: ez a mechanizmus legalább elviekben képes-e az általunk tapasztalt mintázatokat létrehozni. - Szükségszerű, hogy a biológiai rendszerek, például a sejtek leírhatók legyenek kizárólag fizikai összefüggésekkel? - Egyáltalán nem! Az elsődleges érhálózat kialakulása az egyszerűbb jelenségek közé tartozik, az esetek zöme ennél bonyolultabb. Ha találunk egy példát, amelyet sikerül leírnunk, akkor ez igazolja, hogy helyes volt a megközelítés. Persze az összetettebb folyamatokat is szeretnénk megérteni, de annál könnyebb a helyzetünk, minél kevesebb komplex sejtbiológiai tényezőt kell figyelembe venni. Természetesen olyan modellt igyekszünk keresni, amelyik jól leírja a teljes rendszert. A modelljeinkben mikroszkopikus részletekről alkotunk hipotéziseket - például az erek képződésénél feltételezzük, hogyan lépnek a sejtek kölcsönhatásba -, és más skálán nézzük az eredményt. Nem gondolom tehát, hogy a fizika megoldja a biológia problémáit, de szükséges komponens lehet. Az erek kialakulása mellett másik példa a gasztruláció folyamata, ami szintén olyan komplex sejt- és fejlődésbiológiai kérdés, ahol nagy szerep jut a fizikai effektusoknak. Gasztrulációkor - a kezdeti embriófejlődés során - az embrióban kialakulnak a csíralemezek, miközben sejtrétegek áramlanak egymás mellett, végül a sejtek a helyükre kerülnek. Ebben mechanikai tulajdonságokkal rendelkező szövetek vesznek részt, és az áramlásokat erők mozgatják, amelyeket szeretnénk megismerni. Ezeket az erőket persze a sejtek hozzák létre, de pusztán az a tény, hogy fizikai testekről van szó, befolyásolja a lehetséges mozgásformákat egy fejlődő embrióban. Végül, ha meg is ismerjük azokat az erőket, amelyek a folyamatot hajtják, még mindig külön feladatot jelent, hogy meg tudjuk magyarázni a kialakuló áramlási teret. Tehát fizikára mindenképpen szükség van a munkánkban.
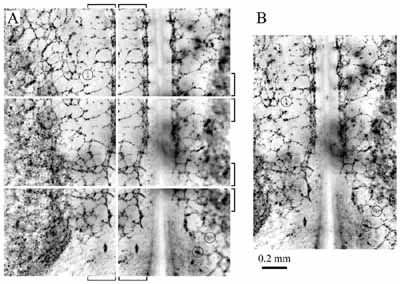
A mikroszkópos képeket egy algoritmus automatikusan mozaikká rendezi a fókuszban lévő részletek megkeresésével - A szövetek felépülésének vizsgálatakor mesterséges szövettenyészetek kialakulását modellezitek, vagy az eredmények vonatkozhatnak a természetes szövetek kialakulására is? - Az egyszerűbb modellek in vitro tenyészetekre vonatkoznak, de az embriógasztrulációt igazából in vivo körülmények között követjük. Van egy rendszerünk, ahol a korai embriót életben tudjuk tartani a mikroszkóp alatt, nagyjából egy napig. Ekkor az egyes sejteket sejtbiológiai módszerekkel megjelöljük, hogy követni tudjuk őket. (Jelölés lehet, ha zölden fluoreszkáló fehérjét termeltetünk velük, vagy valamilyen speciális sejtfelszíni receptor elleni ellenanyagot fluorofórokkal megjelölünk.) Így videomikroszkóppal követhetjük, hogy mi történik a fejlődő embrió sejtjeivel, ami végtelenül izgalmas dolog. - Ez az a videomikroszkóp, amit te fejlesztettél? - Igen, de sokan dolgoztunk rajta. Amikor még doktorandusz voltam az ELTE-n, elkezdtünk fejleszteni egy sejttenyészetek követésére alkalmas videomikroszkópot. Hozzá kell tennem, hogy már 20 évvel ezelőtt is létezett hasonló készülék, de akkor még kevésbé voltak a sejtbiológia előterében a videomikroszkópos vizsgálatok, mert nem létezett kellő számú molekuláris marker, és a technika is nehézkesebb volt. Később, amikor már sokat dolgoztunk sejttenyészetekkel, kimentem posztdoknak az Egyesült Államokba, ahol Charles Little professzor laborjában embriókkal kezdtem foglalkozni, és ott készítettük el a későbbi készüléket. Mikroszkópunk egy automatizált fluoreszcens mikroszkóp, ami kereskedelmi forgalomban is kapható. A mienkben az az egyedülálló, hogy egy embriót életben tarthatunk az objektív alatt, és írtunk egy szoftvert is a kapott képek feldolgozására. A képeket hosszú ideig vesszük fel, sokszor egy napig, több optikai módusban, különféle hullámhosszakon. Ezt az összeállítást így már nem lehet megvásárolni. A legtöbb kutató csak a minta adott helyén nézi a képet három dimenzióban (pl. konfokális mikroszkóppal). Mi egy motorizált asztal segítségével pásztázzuk a szövetet. Így végigkövethetjük az egész érhálózat fejlődését úgy, hogy az egyes sejteket is megfigyeljük. - Ebben az állapotában még nem látszik, hogy madarat vizsgálsz. - Nem, ez egy sejtréteg, amelyben már a gerinchúr és az érszegmensek kezdenek kialakulni, de keringés még nincs, szív sincs. Ezen a szöveten jól lehet követni az erek képződését, mert a sejtréteg szinte kétdimenziósnak tűnik. Részben azért is vizsgálunk madárembriókat, mert ebben a korai stádiumban úgy néznek ki, mint egy nagy palacsinta. Széltében-hosszában több milliméter kiterjedésűek, de a magasságuk csak 300 mikrométer. Tehát a mikroszkópos rendszerünkkel egyrészt meg tudjuk mondani, hogyan alakítják ki az egyes sejtek a korai érhálózatot, másrészt fel tudjuk térképezni azokat a nagy szövetáramlásokat, amelyek a gasztrulációs időszak körül jellemzők az embriókban. Amikor rendelkezésünkre áll a rengeteg adat, akkor megpróbáljuk megérteni, miért pont olyan az áramlási kép, vagy miért pont olyan a sejtek eloszlása, mint ahogy látjuk. Az áramlási tér esetében ezt biomechanikai modellel közelítjük, és eredményeink alapján arra gondolunk, hogy a korai embrionális szövet ún. nem-newtoni folyadékként viselkedik. Szilárd testek és folyadékok esetében nyíráskor mechanikai feszültség lép fel, de egy közönséges folyadékban, például vízben ez a feszültség a nyírás megszűnésekor azonnal eltűnik. A sejtek is hatékonyan tudják csökkenteni a kialakuló mechanikai feszültséget úgy, hogy odébb mennek és megbontják a sejt-sejt kapcsolatokat. Ehhez azonban több időre van szükségük, mint a vízmolekuláknak. Sejtaggregátumon végzett mérések arra utalnak, hogy a sejtaggregátumok (és a szövetek) nem-newtoni folyadéknak tekinthetők, olyasminek, mint egyes polimeroldatok. A természetes szövetekkel kapcsolatban van egy tervünk is. Nagyon sokan dolgoznak a mesterséges erek létrehozásán. In vitro körülmények között, egy gélben endotél sejteket növesztenek, amiből megfelelő körülmények között ki tud alakulni egy csőszerű képződmény. Szeretnénk, ha ez a folyamat önfenntartó lenne és olyan állapotba tudnák hozni a sejteket, mint a valódi ereknél: simaizom is körülvenné őket és meglenne a jó extracelluláris alaphártya. - Vannak még orvosi alkalmazási lehetőségek? - Persze, rengeteg
kutató vizsgálja, hogyan reagálnak az endotél sejtek a környező kemoattraktáns
faktorokra. Mi például együttműködünk egy kollégával a Scripps Intézetből,
aki azzal foglalkozik, hogyan lehet megakadályozni az angiogenezist, vagyis
a tumorsejtek érellátását. Ha egy tumorsejt VEGF-et termel, maga felé vonzza
az érképző, endotél sejteket. David Cheresh professzor vegyülete az endotélsejtek
válaszképességét csökkenti. Tehát gátolja azt a képességüket, hogy VEGF
hatására a tumorhoz vándoroljanak, és ezzel jelentős mértékben hátráltatni
lehet a rosszindulatú tumorok növekedését. Egészséges esetben az erek kialakulásának
egyik lépése, hogy az endotél sejteken jelen lévő két molekula - a mátrix
metalloproteáz 2 (mmp2) enzim és az avß3
integrin - kölcsönhatásba lépjen. Cheresh professzor molekulája ezt a folyamatot
gátolja, mi pedig arra próbálunk rájönni, hogyan hat ez a vegyület és mennyire
változtatja meg az aktívan mozgó endotélsejtek motilitását.
Az ELTE-n több kollégámmal in vitro körülmények között vizsgáltuk különböző agydaganatokból izolált sejtek motilitását taxol és sugárkezelés esetén. Az agydaganatoknál sokszor kialakul egy diffúz "sejtköd" rosszindulatú sejtekből. Sajnos ilyenkor a tumor nemcsak a véráramon keresztül képez áttétet, hanem az agyszövetben is terjed. Mi tumorból izolált sejteken alkalmaztunk taxolkezelést és olyan dózisú besugárzásokat, amit terápiában is használnak. Elég nagy meglepetésünkre az derült ki, hogy ezek a szubletális dózisok sok esetben megnövelik a sejtek motilitását, valószínűleg azért, mert stresszválaszt váltanak ki. Azt találtuk, hogy ha egyszerre adunk taxolt és sugárkezelést, akkor ezek egymás motilitást serkentő hatását csökkentik, tehát optimalizálni kell a kettő arányát. Azt korában is lehetett tudni, hogy a taxol radioszenzitizáló szer, de talán ennek a molekulának a működéséhez is közelebb jutottunk. - Dolgoztál az ELTE-n, a Kansasi Egyetemen, most megint itthon vagy, aztán terveid szerint újra Kansasban… Mi a véleményed, külföldön lehet/kell karriert csinálni vagy inkább itthon? - Ebben nem lehet
tanácsot adni senkinek. Az biztos, hogy az Egyesült Államokban hatalmas,
komoly géppark van, lézerdisszekciós mikroszkóp és minden, amit az ember
el tud képzelni. A kutatási támogatás is sokkal nagyobb, mint Magyarországon.
Kint hatalmas a piac, évente több száz, előre nem "leosztott" állást hirdetnek
meg. Nagy a "motilitás", minden évben elmennek emberek és új doktorandszok,
posztdokok jöhetnek.
- Mindig ugyanahhoz a csoporthoz jársz? - Igen, Little professzor csoportjában dolgoztam, „research assistant professor” státuszban. Ez a gyakornoki státusz után következik, az ember már jogosult NIH-pályázat beadására. - Tavaly el is nyertél egy 1,3 millió dollár költségvetésű pályázatot. - Igen, jó fejlemény,
hogy most már vannak saját pályázataim, mert ott is előbb-utóbb függetlenednem
kell. Ha az ember egyetemi professzor szeretne lenni, akkor elvárják, hogy
felmutasson néhány korábban elnyert, hasonló pályázatot. Az NIH a kutatásfinanszírozás
egyik "egysége".
- Mikor kezdtél el kutatni? - Nagyon korán. Már
elsős egyetemista koromban elmentem Vicsek Tamás professzor egyik speciális
előadására, le is vizsgáztam, és utólag derült ki, hogy elsőéves vagyok.
Ekkor felajánlotta, hogy dolgozzam vele, aminek nagyon örültem. Remek témavezető
volt. Általa jutottam el különböző helyekre is: Ben Jacob professzorhoz
Tel-Avivba, Stanley professzorhoz Bostonba, aztán volt diákjához, Barabási
Albert-Lászlóhoz. A kutatás sokkal jobban érdekelt, mint az egyetemre járás.
- Fizikusként nem riasztottak vissza a biológiai kísérletek? - Nem, mert a feladatok fokozatosan jöttek. Előbb a baktériumok, aztán a sejtek, majd az embriók. A kísérletek során sokat pipettáztam és megtanultam kiszedni a tojásból az embriót. - Külföldön és itthon is neves laboratóriumokban dolgoztál. Hogy látod, mi tesz sikeressé egy labort? - Inkább csak általánosságokat tudok mondani. Az a nagy különbség, hogy más a „kultúra” a fizikusoknál és más a biológusoknál. A fizikusoknál nagyobb a szabadság, nincsenek Journal Club-összejövetelek, de akikkel én találkoztam, azok inkább elméleti emberek, nincsenek is annyira ráutalva az együttműködésre. Egy jó biológuscsoportban pedig - úgy gondolom - erre szükség van: az ember pontosan tudja, mit csinál a mellette ülő. Az is fontos, hogy mindenkinek jól definiált projektje legyen, amit végigvihet. Nyilván mindenütt az a meghatározó, hogy milyenek az emberek: ahol lelkesek és pezseg az élet, ott jó dolgozni. - Mikor fordultál a biológia felé? Középiskolában inkább a matematika és a fizika érdekelhetett, hiszen a KÖMAL versenyein meglehetősen jó eredményekkel szerepeltél. - Gimnázium után azért nem mentem biológusnak, mert visszariasztott, hogy meg kell tanulni a törzsfejlődési, a taxonómiai táblákat, de a sejtbiológia, a molekuláris biológia, amit távolról szemléltem, akkor is érdekelt. Az egyetemen fordultam a biológiai mintázatok felé, amikor Vicsek Tamás csoportjába kerültem. - Mit "tud" a miskolci Földes Ferenc Gimnázium, hogy annyi diákja nyer tanulmányi versenyeket? - Az én időmben Kálmán László tanár úr volt az igazgató, aki matektanár. Ő értékelte a versenyeken való részvételt és elég liberális volt ahhoz, hogy különböző privilégiumaink lehessenek: feljárhattunk szakkörre Pestre, így azokon a napokon nem kellett iskolába járnunk, de ehhez természetesen teljesíteni is kellett. Meg aztán voltak speciális matematikatagozatos osztályok, ahol a tanárok szintén ebben a szellemben dolgoztak. De a többi tanárom is nagyon jó volt, például a magyar- és a történelemtanár, csak én inkább a fizika iránt érdeklődtem. - Mikor szoktál maratont futni? - Csak félmaratont. Maratonit még nem futottam, a félmaraton a leghosszabb, amit be mertem vállalni. Gimnáziumban nem szerettem a testnevelést, csak a kollégiumban kezdtük el a futást, a feleségemmel meg a barátaimmal, azóta ezt többé-kevésbé rendszeresen űzzük. A maratonihoz komolyan kell készülni, de egy félmaratonra szerintem bárki képes. Az interjút készítette: SIMON ÁGNES
|
 -
Alapvető különbség, hogy szervetlen mintázatok (kristályok, fraktálok)
esetén nagyjából ismerjük azokat a mikroszkopikus szabályokat, amelyek
a mintázatot létrehozzák. A mintázatok kialakulását egy reakció-diffúziós
rendszerben sem ismerjük teljesen, de ott legalább azt tudjuk, hogy a molekulák
mozgása és reakciói milyen szabályoknak engedelmeskednek, tehát az alapokat
ismerjük. A sejtek viszont már önmagukban roppant összetettek, ezért arra
is hipotéziseket kell tennünk, hogy egy bizonyos sejt különböző külső környezetekben
hogyan viselkedik. Így az élő szervezetben fellépő kollektív rendszerek
vizsgálata sokkal gyengébb alapokon áll, mint a fizikai rendszerek tanulmányozása.
-
Alapvető különbség, hogy szervetlen mintázatok (kristályok, fraktálok)
esetén nagyjából ismerjük azokat a mikroszkopikus szabályokat, amelyek
a mintázatot létrehozzák. A mintázatok kialakulását egy reakció-diffúziós
rendszerben sem ismerjük teljesen, de ott legalább azt tudjuk, hogy a molekulák
mozgása és reakciói milyen szabályoknak engedelmeskednek, tehát az alapokat
ismerjük. A sejtek viszont már önmagukban roppant összetettek, ezért arra
is hipotéziseket kell tennünk, hogy egy bizonyos sejt különböző külső környezetekben
hogyan viselkedik. Így az élő szervezetben fellépő kollektív rendszerek
vizsgálata sokkal gyengébb alapokon áll, mint a fizikai rendszerek tanulmányozása.