A kromoszómák sejtbéli rendje és a mesterséges kromoszóma
A genomprogramok sikere a különbözõ fajok DNS-bázissorrendjének megállapítása alapján megjósolható, hogy az elkövetkezendõ évtizedek meghatározó tudománya a genetika, illetve az azt kiszolgáló molekuláris biológia, bioinformatika lesz. A megismert DNS-szekvenciák értelmezése, a kódolt gének és a nem kódoló DNS funkciójának, a gének, valamint a szabályzó szakaszok kölcsönhatásainak kiderítése az emberiség történetének olyan mérföldköve lesz, ami az emberi faj fejlõdése és fennmaradása szempontjából is meghatározó lehet. Elképzelhetõ, hogy huszonöt, ötven év múlva úgy tekintünk vissza az átöröklés alapvetõ szabályainak újrafelfedezése óta eltelt száz év lélegzetelállító történéseire, mint a genetikai robbanás csendes elõjátékára.
A befejezéshez közeledõ humángenom-program (az emberi DNS bázissorrendjének meghatározása) jól szemlélteti a feladat nagyságát. Ha a 33,5 milliárdnyi betûvel leírható bázissorrendet az 1800-as évek elejérõl származó Károlyi Bibliához hasonlítjuk, akkor az emberi DNS-t leíró szöveg ezerszerese a 863 oldalas Ótestamentumnak. A genetikusok, molekuláris biológusok és informatikai szakemberek elõtt álló munka nem kevesebb, mint e szöveg értelmezése, a funkcióval rendelkezõ és szerkezeti DNS-szakaszok meghatározása, és azok feladatának kiderítése (funkcionális genomika). Ennek egyik várható eredményeként könnyen megjósolható, hogy a kromoszómaszerkezet és a -mûködés, valamint a sejtmag mûködésének másfél évszázados titka hamarosan csak egy-egy tényszerû fejezet lesz a genetika 21. századi forradalmának történetében.
 |
| 1. ábra. A kromoszóma
elektronmikroszkópos képe. Ennek a kromo- szómának mindkét kro- matidája kb. 250 millió bázispárnyi DNS-t hordoz. |
A kromoszómakutatás története méltó e zseniális kis szerkezetekhez, egyáltalán nem szokványos, bõvelkedik a furcsaságokban. A kromoszómakutatás fejlõdése elválaszthatatlan a genetikáétól, így már csak emiatt is érdemes rövid történeti visszatekintést tenni e területen. A kromoszómák történelmének furcsa fintora, hogy az a svájci botanikus, Karl Wilhelm von Nägeli, aki 1842-ben huszonöt évesen elsõként leírta és lerajzolta a kromoszómákat (melyeket citoblasztoknak nevezett el), két évtizeddel késõbb, a müncheni egyetem professzoraként elutasította egy kézirat közlését, amely nem kevesebbet írt le, mint a kromoszómális öröklõdés szabályait. E tanulmány szerzõje a brnói matematikus szerzetes, GregorJohann Mendel volt, akinek szabályait immár egy évszázada mendeli öröklõdésnek nevezzük. (Nägeli, aki korának kiemelkedõ és elismert tudósa volt, eme tévedéséért nagy árat fizetett, a tudománytörténészek többségének sommás értékelése szerint ugyanis többet ártott, mint használt a biológiának.) A kromoszómákat felfedezésüket követõen csaknem negyven éven át gyakorlatilag figyelemre sem méltatták, mígnem Walter Flemming, Eduard Strasburger, Edouard van Beneden és Wilhelm Roux munkáikban a titokzatos fonalaknak már hipotetikus szerepet tulajdonítottak az öröklésben. 1871-ben Johann Friedrich Miescher izolált egy anyagot a fehérvérsejtekbõl (valójában nukleinsav és fehérjék keverékét), amelyet nuclein-nek nevezett el. (A nukleinsav kifejezés 1899-bõl Richard Altmanntól származik.) 1882-ben Walther Flemming a sejtmag állományát chromatin-nak keresztelte el, pontosan leírta a testi sejtek osztódását, amelynek a mitosis nevet adta. Nevéhez fûzõdik az elsõ kromoszómaszám meghatározása és a kromoszómák (kromatidák) hosszanti szétválásának pontos leírása.
A citológiai anilinfestékek megjelenését követõen a kissé megkésett keresztelõre is sor került. Negyvenhat évvel felfedezésük után a citoblasztok, a titokzatos fonalak erõs festõdésük alapján a német Heinrich Wilhelm Gottfried Waldeyertõl a kromoszóma nevet kapták (a görög HROMA=színes, SOMA=test összevonásából).
Waldeyer a keresztapja a kromoszómák legfontosabb szerkezeti elemének, a centromernek is, ahová a sejtosztódások során, a kromoszómák (kromatidák) mozgását biztosító húzófonalak tapadnak. Kis túlzással, ennél többet ma sem tudunk a centromerrõl. Nem tudjuk, hogy milyen DNS-szakaszok szükségesek a centromer kialakulásához, vannak-e olyan DNS-szakaszok, amik feltétlenül szükségesek a centromer kialakulásához. Emlõsök sejthibridjein végzett vizsgálatok eredményei alapján annyi bizonyos, hogy az emlõsfajok teljesen eltérõ DNS-ébõl (és a hasonló, de különbözõ fehérjékbõl) felépülõ centromerjei egymás sejtjeiben mûködõképesek. Vagyis az eltérõ molekuláris felépítés ellenére idegen környezetben is képesek feladatuk ellátására.
1902-ben, két évvel a Mendel-szabályok újrafelfedezését követõen, az amerikai Walter Stanborough Sutton kísérletei alapján megszületett a kromoszómaelmélet, miszerint a normális fejlõdéshez teljes kromoszómaállomány szükséges, az egyes kromoszómák eltérõ genetikai tartalommal rendelkeznek és az egyes kromoszómapárok tagjai függetlenül (véletlenszerûen) jutnak az ivarsejtekbe. Sutton, munkájának gyümölcseként, 1906-ban doktori fokozatot szerzett, és ugyanebben az évben az öröklõdés fiatal tudománya William Batesontól a genetika nevet kapta.
A mendeli genetika szabályainak újrafelfedezése gátszakadásszerû felfedezésáradatot idézett elõ, melyben három évtizeden keresztül a kromoszómák (és fõként az ecetmuslica) központi szerepet játszottak. 1927-ben Theophilus ShickelPainter egy kromoszómaszakasz hiányából eredõ genetikai következmények tanulmányozása révén elsõként határozta meg egy gén helyét az emlõskromoszómán. Theodosius Dobzhansky, Theophilus Shickel Painter és Hermann Joseph Muller (1932) munkássága nyomán bebizonyosodott, hogy a gének sorrendje meghatározott az egyes kromoszómákon, azaz a kromoszómatérkép (citológiai térkép) és a géntérkép (genetikai térkép) azonos. Késõbb, 1946-ban Muller a röntgensugarak mutagén hatásának felismeréséért Nobel-díjat kapott. Ugyanez az elismerés jutott 1933-ban Thomas Hunt Morgannak, aki a kromoszómák öröklésben betöltött alapvetõ szerepét fedezte fel.
Ezt követte a baktérium- és fággenetika születése és a molekuláris genetika meghatározó korszaka, ami olyan diadalmenetnek bizonyult, hogy ennek akár vázlatos ismertetésére sincsen mód e helyen. A téma önálló, külön írást érdemel. E korszak felfedezéseinek jelentõségét három önkényesen kiemelt mérföldkõ is szemlélteti: 1952-ben Alfred Day Hershey és Martha Chase bizonyította, hogy a DNS örökítõanyagot tartalmaz, 1953-ban James Dewey Watson és Francis Harry Compton Crick leírta a DNS szerkezetét és elõrevetítette a szerkezetbõl adódó genetikai funkciót, majd 1961-ben Marshall Warren Nirenberg és Har Gobind Khorana munkája nyomán megkezdõdött a genetikai kód feltörése. Megállapították, hogy a DNS-t felépítõ bázisok hármas csoportjai (tripletek) egy-egy aminosavat határoznak meg (kódolnak). A DNS bázissorrendjébõl (szekvencia) így leolvashatóvá vált, hogy egy-egy DNS-szakasz milyen fehérjéket kódol.
1956-ban Joe Hin Tjio és Albert Levan kétséget kizáróan bebizonyította, hogy az ember testi sejtjeinek kromoszómaszáma 46. Ezzel megkezdõdött a humán klinikai citogenetika hajnala, a kromoszómaszám-rendellenességek genetikai következményeinek feltérképezése. 1957-ben egy évvel a DNS szemikonzervatív replikációjának bizonyítása elõtt, amely Matthew Stanley Meselson és Franklin William Stahl nevéhez fûzõdik James Herbert Taylor, Philip Sargent Woods és Walter L. Hughes radioaktív timidin beépítésével igazolta ugyanezt a kromoszómáknál. Az ötvenes évek végét számos, meghatározó jelentõségû felismerés fémjelzi: az ivari meghatározottság kromoszomális alapjainak felderítése (Charles Edmund Ford, Patricia A. Jacobs és Joe Hin Tjio), egy számfeletti kromoszóma felfedezése a Down-szindrómás (21-es triszómia) gyermekek sejtmagjában (Jerome LeJeune, Martha Gautier és Raymond Alexandre Turpin). Ezt hamarosan követte az ivari kromoszómák rendellenes számából adódó fejlõdési rendellenességek azonosítása. Például az egyik X-kromoszóma hiányának felfedezése a Turner-szindrómában (X0) szenvedõ nõk sejtjeiben, vagy egy számfeletti X-kromoszóma jelenléte a Klinefelter-szindrómás (XXY) betegeknél.
A hetvenes években a kromoszómák egyedi sávfestésének kidolgozásával lehetõvé vált, hogy az egyes kromoszómákat teljes biztonsággal azonosítsák, amely napjainkra a fluoreszcens DNS-hibridizálással és a számítógépes képanalizáló rendszerek kifejlesztésével kiegészülve klinikai diagnosztikai iparággá vált.
Az 1960-as években kezdõdött a kromoszómák elektronmikroszkópos szerkezetvizsgálata, ami a hetvenes években biokémiai módszerekkel gazdagodva, a kromoszómaváz (scaffold) modelljének megszületését eredményezte. Eszerint a kromoszómákon belül egy fehérjékbõl álló folytonos váz szervezi a DNS-t a kromatidák mentén hatalmas, körkörös hurkokba. A késõbbi, részletes vizsgálatok eredményei azonban kérdésessé teszik az ilyen folytonos fehérjeváz létét. Sokkal valószínûbb, hogy a kromoszómák vázát fehérjeDNS-részecskék halmaza adja, azaz a kromoszómális DNS bizonyos szakaszai szerkezetépítõ szerepet töltenek be. Ugyanezen vizsgálatok tanúsága szerint a kromoszómák szerkezetileg legellenállóbb szakasza a centromer (2. ábra).

2. ábra. Egy részlegesen fehérjétlenített
kromoszóma elektronmikroszkópos
képe. A kromoszóma alakját kirajzolja a fehérjeDNS-részecskék
eloszlása,
de a kromoszóma egyetlen, magasabb rendû szervezettséget
mutató szakasza, a centromer két gömböcske
formájában megmarad.
Ennek alapján feltételezhetjük, hogy a centromer a sejtosztódások során betöltött alapvetõ feladatán túl a kromoszómák szerkezeti szervezõdésében is szerepet játszhat.
A már említett mérföldkövek a 20. század közepén a DNS-szerkezet és a genetikai kód megfejtésének genetikai következménye: a molekuláris genetika, a molekuláris biológia korszaka. E napjainkig tartó idõszak ipari csúcsteljesítménye a humángenomprogram sikere.
A humángenom-program eredményeit ünnepelve a gyakran túlzó várakozások miatt nem árt megjegyezni, hogy egyelõre távol vagyunk valamennyi génünk azonosításától. Aki már holnapra várja a genetikai csodát, annak érdemes megszívlelni Sir John Maddox (aki 23 éven keresztül volt a világ egyik legtekintélyesebb természettudományi folyóiratának, a Nature-nak fõszerkesztõje) figyelmeztetését, miszerint a körülbelül 100 ezer génünk pontos funkciójának megismerése akár tízszer annyi idõbe is telhet, mint génjeink azonosítása.
 |
| 3. ábra. A részlegesen fehér-
jétlenített emlõssejt magjából kiáramló DNS mennyisége jól szemlélteti a sejtmagban található DNS tömegét |
A sejtmag finomszerkezetét az egyre fejlettebb fény- és elektronmikroszkópos módszerekkel vizsgálva kiderült, hogy a sejtmagban rendezetlen, lazább-tömörebb (eukromatin-heterokromatin) területek figyelhetõk meg. A sejtmagon belül a riboszóma RNS-gyárak a sejtmagvacskák (nukleoluszok) voltak a funkcionális szempontból egyedül elkülöníthetõ sejtmagi területek. Az 1980-as évek közepéig a sejtmagot többnyire olyan sûrû, szerkezet nélküli spagettilevesnek tartották, amiben hosszú DNS-fonalakhoz kötõdõ fehérjék rendezetlen halmazát a maghártya tartja össze. A sejtmagi káosz valódisága mindinkább kérdésessé vált elméleti megfontolások és kísérleti eredmények alapján egyaránt. A tavaly Nobel-díjjal kitüntetett Günter Blobel 1985-ben megalkotta a gene gating hipotézist, miszerint a sejtmaghártya kapuinak rendezett, nem véletlenszerû eloszlása a sejtmag funkcionális rendjének megfelelõ, a sejtmag térbeli mûködési egységei tükrözõdnek a sejtmaghártya felszínén. Az ecetmuslica óriáskromoszómáinak, illetve az egér humán sejthibridek kromoszómáinak sejtmagi helyzetét vizsgálva kezdett kialakulni az a nézet, hogy az egyes kromoszómák meghatározott sejtmagi területeket foglalnak el. Erre a rendre 1985-ben sikerült kísérleti bizonyítékot is szolgáltatnunk a kevés kromoszómaszámú (13 kromoszómát hordozó) erszényes patkány laboratóriumi körülmények között tenyésztett sejtjein végzett munkánk során. A kromoszómák sejtmagi helyzetét a centromerek immunofluoreszcenciás festésével vizsgálva kiderült, hogy az egyes kromoszómák sajátos rendben találhatók. Ez a rend kétszintû.
1. Az egyes kromoszómapárok anyai és apai tagja (homológ kromoszómák) szorosan egymás közelében találhatók. Ez a növényi és a rovarsejteknél jól ismert jelenség a homológ kromoszómák asszociációja.
2. A nem homológ kromoszómák két vagy több szimmetrikus csoportban, rendezetten figyelhetõk meg a sejtmagban (4. ábra).

4. ábra. Az erszényes patkány kromoszómáinak
helyzete a sejtmagban.
Az egyes kromoszómák centromerjeinek helyzetét
a világos pontok mutatják (a).
Ugyanerrõl a sejtmagról fáziskontraszt-mikorszkóppal
készített kép (b).
E szimmetrikus rend nem csak az erszényes patkány kromoszómáinak sajátossága. Késõbbi vizsgálatok tanúsága szerint a szimmetrikus nem homológ kromoszómacsoportok léte igazolható volt a 40 kromoszómát hordozó egér sejtjeiben is.
A sejtosztódások során a kromoszómák
gyûrû alakban rendezõdnek, a kromatidák szétválását
követõen a két kromatidagyûrû az utódsejtekbe
kerül, és létrehozzák a két új
sejtmagot. Feltételezhetõ, hogy az egyes kromoszómák
kromoszómagyûrûn belüli helyzete megõrzõdik
a sejtmagban is. Ebbõl következõen az egyes kromoszómák
egyedi festését biztosító hibridizációs
próbákkal elvileg elkészíthetõ lenne
a sejtmagtérkép. Azaz, ha létezik egy meghatározott,
akár sejttípusra vagy egyedre jellemzõ topológiai
viszony az egyes homológ és nem homológ kromoszómák
között, ez egyértelmûen megállapítható
(5.
ábra).
 |
 |
| 5. ábra. Emberi "kromoszómagyûrû",
amelyben a 8-as és 14-es számú kromoszóma-
pár helyzete látható (a). A 8-as anyai és apai kromoszómák helyzete mutatja, hogy az egyes kromoszómák gyûrûn belüli helye nem változik az anafázisban sem (b) |
|
Az ilyen vizsgálat választ adhatna olyan kérdésekre is, hogy például bizonyos megbetegedésekben, ahol a nem homológ kromoszómák között kromoszómadarabok cseréjére kerül sor (transzlokáció), a kromoszómák sejtmagtérképen belüli helyzete szerepet játszik-e a kór kialakulásában vagy sem.
Ma még nem tudjuk, hogy szerkezeti szempontból mi biztosítja a nem homológ kromoszómák megfigyelt rendjét, hogy az eltérõ feladatot ellátó sejtekben azonos-e ez a rend, s hogy mi a szerepe a kromoszómák eme rendjének a sejtmag mûködésében?
A kérdéseket még hosszan sorolhatnánk, mint
ahogyan azt a sok száz molekuláris biológiai, biokémiai,
genetikai munkát is, melyek eredménye alapján már
napjainkban is vitathatatlan, hogy a sejtmag az eddig ismert legbonyolultabb
nanogépezet. Napról napra többet tudunk e gépezet
önálló, finoman szabályozott részegységeinek
(kompartmentjeinek) összehangolt mûködésérõl.
Bizonyított, hogy a közös feladatot ellátó
géncsoportok összehangolt szabályozás alatt állnak,
a gének egymáshoz viszonyított fizikai közelsége
vagy távolsága befolyásolja a géntermékek
képzõdését. A gének termékének
utómunkálatai a sejtmagon belül elkülönült
RNS-mûhelyekben történik, a kész hírvivõ
RNS-ek citoplazmába szállítása meghatározott
sejtmagi csatornákon keresztül zajlik
(6. ábra).
 |
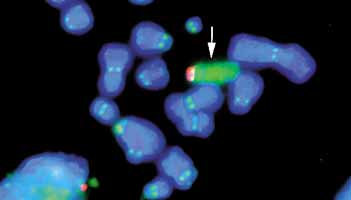 |
| 6. ábra. Géntermék szállítása
sejt-
magi csatornán keresztül. A kék színû sejtmagban a két kromoszó- ma helyzetét a piros foltok jelzik. Az egyik kromoszómáról képzõ- dött géntermék (hírvivõ RNS) szál- lítási útvonalát a zöld szín mutatja. (Dr. Szeles Anna felvétele, Karo- linska Intézet, Stockholm) |
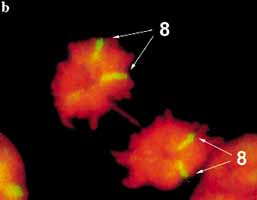
7. ábra. Emberi mesterséges kromoszóma (zöld-piros)
természetes kromoszómák társaságában. A mesterséges kormoszóma festõdése az eltérõ DNS-összetételt jelzi. A kromoszómákon látható kettõs pöttyök a centrome- reket jelzik. |
A citoplazma és a sejtmag közötti, kétirányú jelátvitel, makromolekulák, fehérjék forgalmának egyre pontosabb megismerése stb., bár újabb és újabb kérdéseket vet fel, joggal remélhetjük a titok megfejtését.
A kromoszómatörténet jelene: a mesterséges kromoszómák létrehozása. A kilencvenes években kiderült, hogy az emlõsök sejtjei bizonyos körülmények között képesek új kromoszómákat létrehozni. Ez a folyamat kísérleti úton bekapcsolható és ezzel lehetségessé vált a tetszés szerinti genetikai tartalmú, célirányos kromoszómaépítés élõ sejtekben. Ennek legfrissebb eredménye a mûködõképes emberi mesterséges kromoszómák elõállítása (7. ábra).
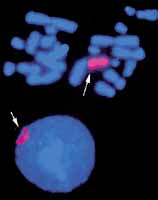 |
| 8. ábra. A mesterséges kro-
moszóma (piros) helyzete a sejtmagban |
A mesterséges kromoszómák létrehozása során az egyik elvi bizonytalansági tényezõ is a sejtmagi renddel volt kapcsolatos. Nevezetesen, ha létezik egy szigorú sejtmagi rend, akkor hogyan illeszthetõ be egy mesterséges kromoszóma ebbe a rendbe anélkül, hogy ott mûködésbeli zavarokat okozna? Ma már többé-kevésbé választ tudunk adni erre a kérdésre. Amennyiben a mesterséges kromoszóma olyan domináns jelzõgént hordoz, amely terméke biztosítja a sejt túlélését, például egy bizonyos mérgezõ anyagot tartalmazó táptalajban is, akkor a sejtmag rákényszerül a mesterséges kromoszóma befogadására. Citológiai vizsgálatok tanúsága szerint ez a túlélést biztosító mesterséges kromoszóma többnyire a sejtmag perifériáján figyelhetõ meg (8. ábra).
Lehetséges, hogy a sejtmag a mûködéséhez szükséges rendet úgy védi meg, hogy a kényszerûen befogadott idegen jövevényt a peremvidéken tartja.
A rend hálója enged,
Majd újra összezárul.
Radnóti Miklós: Talán