Az evolúcióbiológia horizontjai
Kissé talán rendhagyó írásomban
nem egyetlen tudományos kérdéssel foglalkozom, hanem
egy közelmúltban lezajlott konferenciáról tudósítok.
A beszámoló az European Society for Evolutionary Biology
(ESEB) nyolcadik, 2001. augusztus 2025. között Dániában
megrendezett kongresszusán elhangzott elõadások alapján
készült.
Augusztus vége a dániai Aarhusban. Hétszáz biológus zsongása tölti meg az egyetem folyosóit: rég nem látott ismerõsök, újonnan jött fiatalok. Az elsõ este a híreké. Beszélgetés közben a poszterek is felkerülnek az állványokra. A gyors üdvözlések lassan kötetlen, lelkes vitákká mélyülnek. Megkezdõdött az Európai Evolúcióbiológiai Társaság kétévenkénti kongresszusa.
Híven beszámolni egy ilyen eseményrõl bizonyosan nem lehet, hiszen jó háromszáz elõadás hangzott el az öt párhuzamosan folyó szekcióban. A kiragadott témák tehát egy szükségképpen szubjektív válogatást jelentenek.
A molekuláris biológia robbanásszerû fejlõdése az evolúciógenetikában is soha nem látott lehetõségeket nyitott meg. A különbözõ fajok egyes génjeinekösszehasonlítása régóta bevett eszköze annak, hogy felmérjük a fajok rokonsági viszonyait, s génszekvenciáik alapján molekuláris törzsfákat szerkesszünk. Egy gén vizsgálata során azonban nehéz azt megállapítani, hogy a szekvenciában bekövetkezett változás dacára a kódolt fehérje ugyanúgy mûködik-e, mint korábban (valószínûleg a legtöbb különbség ilyen semleges variációnak felel meg), vagy az eltérõ szekvencia eltérõ funkciót takar, s mint ilyen, adaptív evolúció eredménye. A gyors szekvenáló eljárások terjedésével teljes génkészletek (vagy legalábbis számos gén) összehasonlítására is lehetõség nyílik. A teljes emberi génkészlet szekvenálása a közelmúltban készült el, de több más élõlény, így a genetika kedvenc Drosophilájának teljes genomszekvenciája is ismert már. Andrew Clark (USA) elõadásában többek között arról esett szó, hogyan azonosíthatjuk az evolúció során különösen gyorsan változó géneket egy-egy evolúciós leszármazási vonalon belül. Ezek nyilván magukban foglalják az egyes élõlényeket kizárólagosan jellemzõ adaptált tulajdonságok génjeit is. A gyorsan evolválódó gének részletesebb vizsgálata végre megválaszolhatja a genetika egyik nagy kérdését: pontosan mely gének teszik az egeret egérré?
Molekuláktól a fajokig és tovább. Roger Butlin (Nagy-Britannia) és munkatársai a genomszintûösszehasonlítás egy viszonylag durvább módszerét használták egy faj születése körüli részletek felderítésére. A Littorina saxatilis nevû csiga két, fõként a ház formájában eltérõ változata a tengerparti árapályzóna magasabb (hullámoktól védettebb), illetve alacsonyabb (ragadozóktól védettebb) sávjában él, viszonylag elkülönülten. A korábbi vizsgálatok szerint a két változat jó úton halad a külön fajjá válás felé. Butlinék a két változat DNS-ét olyan enzimekkel (ún. restrikciós endonukleázokkal) vágták darabokra, amelyek szigorúan meghatározott rövid szekvenciáknál hasítanak (hasítóhelyek a genomban mindenfelé találhatók). A keletkezõ darabok méreteinek alapján a hasítóhelyek megléte vagy hiánya vethetõ össze: természetesen ez jóval kevesebb, mint a változatok teljes genomszekvenciájának összehasonlítása, de azért ez mégsem csak egyetlen génen alapuló, hanem az egész génkészletre kiterjedõ módszer. Ennek segítségével megállapították, hogy a gének mintegy 4 százaléka erõsebben különbözik a térbeli elkülönülés alapján várhatónál. Természetesen itt nem tudjuk, mely génekrõl van szó, mert nem tudjuk, hogy melyik hasítóhely melyik génhez tartozik. Az elõadást mégis nagy érdeklõdéssel hallgatom. Miért? A fajképzõdés hagyományos elmélete szerint a fajok szétválása hosszú idõ alatt történik, s eközben nagyon sok kisebb-nagyobb véletlen genetikai eltérés halmozódik fel. Az ezzel szemben álló, úgynevezett adaptív speciáció elmélete szerint viszont a fajkeletkezést a természetes szelekció hajtja, a fajok szétválása a különbözõ környezetekhez való alkalmazkodás eredménye. Az elsõ modell alapján sok kis genetikai különbséget várhatunk a születõ fajok között, az utóbbi szerint viszont eleinte csak néhány, az adaptációban fontos gén válik erõsen különbözõvé. Butlinék eredménye hogy a gének egy kis százalékában kiugróan nagy különbségek vannak az utóbbi, adaptív fajkeletkezést támasztja alá.
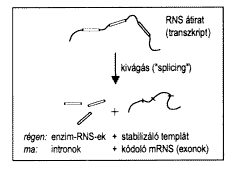
1. ábra. Az intronok lehetséges evolúciós eredete. Az intronok (téglalapok) az exonok (vonalszakaszok) közé beékelt szekvenciák, amelyek az RNS-átiratból a fehérjeszintézis elôtt kivágódnak. Ma az exonok játsszák a központi szerepet, mivel ezek kódolják a fehérjeenzimek aminosavsorrendjét. Régen azonban az intronok lehettek a lényegesek, mert saját enzimaktivitással rendelkeztek.
Jóformán véletlenül kukkantok be a DNS fehérjéket nem kódoló szakaszainakevolúciójával foglalkozó szekcióba, de nem bánom meg: D. Jeffares (Dánia) az intronok igen korai eredete mellett hoz fel érveket. Az intronok a géneken belül elszórtan található nem kódoló szekvenciák, amelyeket a génrõl készült RNS-átiratból a fehérjeszintézis elõtt ki kell vágni (1. ábra). Ez persze felesleges bonyodalmat jelent a fehérjeszintézis során, így a céltalannak látszó intronok eredete méltán izgatja az evolúciógenetikusokat. Lehetséges, hogy az intronok pusztán önzõ DNS-darabok, tulajdonképpen molekuláris paraziták, amelyek önmagukat másolva egyre több helyen ékelõdtek be a genomba. Ám az is lehet, hogy az intronok eredete még arra az idõre nyúlik vissza, amikor RNS-molekulák katalizálták a sejt kémiai reakcióit a mai fehérjeenzimek helyett. Egyes intronok RNS-átiratai még ma is rendelkeznek enzimaktivitással. Közülük kiemelkedik az snoRNS (small nucleolar RNS) elnevezésû csoportba tartozó számos intron, melyek a riboszómák összeállításában játszanak szerepet. D. Jeffares és munkatársai úgy képzelik, hogy ezek az intronok már léteztek a fehérjeszintézis hajnalán és aktív RNS-enzimekként mûködtek. Az õket körülvevõ szekvenciák, amelyek az intronok kivágása után visszamaradtak, eleinte talán csak a stabilizátor szerepét töltötték be a peptidszintézisben. Ezek a stabilizáló szekvenciák azután úgy evolválódtak, hogy rajtuk a legmegfelelõbb fehérjéket lehessen összerakni: végeredményben ezekbõl lettek volna a mai exonok, vagyis az aminosavak sorrendjét meghatározó szakaszok. Ha nem is valamennyi mai intron tekinthet vissza hosszabb evolúciós múltra, mint a fehérjéket kódoló exonok, az intronok kivágásának mechanizmusa igen õsi lenne, és az snoRNS intronokat tartalmazó gének termékeiben talán a legrégibb fehérjéket tisztelhetjük.
Mikroorganizmusok a sejten belül
Andrew Read (Nagy-Britannia) plenáris elõadása a malária evolúciójáról vet fel új kérdéseket, sõt orvosi dilemmákat. A malária kórokozója, a Falciparum malariae nevû egysejtû parazita a vörös vértestekben szaporodik, és a fertõzött emberrõl a maláriaszúnyog közvetítésével jut át újabb áldozataiba. Minél gyorsabban szaporodik a kórokozó a szervezeten belül, vagyis minél virulensebb, annál nagyobb számban van jelen a vérben, és így annál valószínûbb, hogy sikeresen jut át a közvetítõ szúnyogba. Vajon mi szab gátat a virulencia növekedésének? Valószínûleg az, hogy a sok kórokozó jelenléte a betegség súlyos formájához és gyors halálhoz vezet, s így a virulensebb törzseknek nincs idejük arra, hogy gazdájuk halála elõtt sok szúnyogot fertõzzenek meg. Ám a maláriás fertõzéseknek csak mintegy 1 százaléka vezet gyors halálhoz. Elegendõ lenne az ilyen alacsony halálozási arány ahhoz, hogy megakadályozza a virulencia növekedését, vagy ha nem, akkor miért nem válik a malária agresszívebbé? A malária a Föld jelentékeny részének rettenetes betegsége, az okozott halálesetek számát évente 1 és 2,7 millió közöttire becsülik. Furcsa, de mégis azt kell kérdeznünk, miért marad aránylag kevéssé virulens ez a kórokozó?
Ezt annál is inkább jó lenne megérteni, nehogy valami olyat tegyünk, ami egy virulensebb maláriatörzs evolúciójához vezetne. Egy ilyen veszélyes lehetõség az oltás bevezetése. A maláriával szemben megbízható természetes immunitás nem alakul ki, és oltással sem lehet ilyet kiváltani. Ismert azonban egy vakcina, amely ha nem is akadályozza meg a fertõzést, de lassítja a kórokozó szaporodását a szervezeten belül. A beoltottak így kevésbé fertõznek, hamarabb felépülnek és kisebb valószínûséggel esnek a betegség áldozatául. Az oltás az egyén szempontjából, és rövid távon a közösség szempontjából is kedvezõ. Ám várható, hogy a kórokozó alkalmazkodni fog az oltás által megváltoztatott körülményekhez. Ha a virulencia jelenlegi szintje felel meg a kórokozó szervezeten belüli optimális szaporodási sebességének, de a szaporodást az oltással lassítjuk, akkor virulensebb törzsek fognak elterjedni, amelyek az oltás dacára a mai szaporodási sebességet produkálják. A kórokozók rövid szaporodási ciklusuknak köszönhetõen gyors alkalmazkodásra képesek, így a virulensebb törzsek elterjedésére a nem túl távoli jövõben számítani kell. Miután ez megtörtént, az oltás folyamatos alkalmazása mellett visszaáll a mai állapot, már ami a megbetegedések és halálozások számát illeti. Az oltást azonban nem lehet abbahagyni, hiszen akkor a virulensebbé vált kórokozó még súlyosabb betegséget okozna. Végeredményben az oltás költségeit hosszú távon kell viselni, haszon pedig hosszú távon semmi sincs. Sõt, az oltás bevezetése után a még beoltatlanok (például újszülöttek) különösen nagy veszélynek lesznek kitéve.
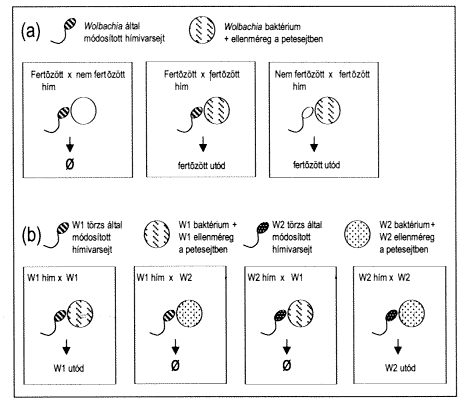
2. ábra. (a) A Wolbachia úgy módosítja a fertôzött hím spermiumát, hogy elpusztítja a keletkezô embriót (elsô panel). Ha viszont a petesejtben jelen van ugyanaz a baktérium, akkor az megvédi az embriót és a keletkezõ fertôzött utód normálisan fejlôdik (második panel). A fertôzés a petesejten átjut az utódba (harmadik panel). (b) A különbözô Wolbachia-törzseket hordozó egyedek nem tudnak utódot nemzeni egymással, mert a különbözô törzsek ellenmérgei nem védenek egymás hatásával szemben.
A Rickettsia-szerû baktériumok közé tartozó Wolbachia szintén a sejteken belül él, számos rovarfajban (de más gerinctelen fajokban is) az ivarszerveket támadja meg elõszeretettel. A fertõzés szisztémás tüneteket nem okoz, de a Wolbachiával fertõzött hímek nem tudják megtermékenyíteni a fertõzés nélküli nõstényeket. A herékben levõ baktérium ugyanis úgy módosítja a hímivarsejteket, hogy a keletkezõ embrió tönkremegy, hacsak nincs a petesejtben szintén jelen a baktérium, mely ott ellenmérget biztosít (2.a ábra). Bár ez a viselkedés kissé bonyolultnak tûnhet, a baktérium szempontjából kétségtelenül elõnyös. A baktérium a petesejteken keresztül átadódik az utódoknak, de a hímivarsejteken keresztül nem. A baktériumot hordozó petesejtek mindig életben maradnak, függetlenül attól, hogy milyen hímivarsejt termékenyíti meg õket. A baktériumot nem hordozó petesejteket azonban a baktérium, ha teheti, a hímivarsejten át elpusztítja. Ez az öldöklés korántsem öncélú: a versenytársak kiiktatásával a megmaradó, baktériumot hordozó utódok jobb életkörülmények közé jutnak. Más szóval, a hímekben levõ, direkt szaporodásra képtelen baktérium azzal segíti a vele azonos géneket hordozó, nõstényekben levõ társainak szaporodását, hogy elpusztítja a fertõzés nélküli utódokat, vagyis a fertõzött utódok leendõ versenytársait.
A Wolbachia egyik törzsével fertõzött hímek nem csak a fertõzés nélküli nõstényeket nem tudják megtermékenyíteni, egy másik Wolbachia törzzsel fertõzötteket sem. A két különbözõ törzzsel fertõzött populáció egyedei így nem képesek egymással szaporodni, vagyis külön fajt alkotnak (2.b ábra). Valóban, találtak olyan közelrokon, de más-más helyen élõ (allopatrikus) rovarfajokat, ahol a reproduktív izolációt kizárólag az eltérõ Wolbachia fertõzés okozza. Ezt roppant egyszerûen lehetett igazolni: ha antibiotikum-kúrával megszüntették a fertõzést, a két faj minden további nélkül keresztezõdött. A Wolbachia okozta reproduktív izoláció talán hozzájárul ahhoz, hogy oly elképesztõen sok rovarfaj létezik!
Vajon szerepet játszhat-e a Wolbachia a földrajzi elkülönülés nélküli (szimpatrikus) fajkeletkezésben is? Diszruptív szelekció (a szélsõséges tulajdonságok elõnye) elég sok ökológiai helyzetben elõfordulhat, s ez egyetlen populációban is szét tudna választani két fajt, ha a reproduktív izolációt biztosítani lehetne valahogy. Reproduktív izoláció nélkül a diszruptív szelekció csak a populáció variabilitását növeli, de nem tud két világosan elkülönülõ típust létrehozni. Martijn Egas és Filipa Vala (Hollandia) vetették fel a gondolatot, hogy a Wolbachia által oly könnyedén megvalósuló reproduktív izoláció segíthetne a diszruptív szelekciónak abban, hogy szimpatrikus fajokat hozzon létre. Az ötletet modellezés követte, s kiderült, hogy a dolgok azért nem mennek olyan simán. A Wolbachia valóban segített a fajok kezdeti szétválásában, de egy idõ után az egyik faj rendre kihalt a modellben. Ha az egyik faj (ökológiai okokból, vagy pusztán véletlenül) kisebb számban van jelen, mint a másik, akkor a ritkább faj egyedei nehezebben találják meg egymást, és gyakrabban próbálkoznak a másik fajjal való illegitim, terméketlen párosodással. Emiatt a ritkább fajban kevesebb utód születik, ami a létszám további csökkenéséhez, végül kihaláshoz vezet. Wolbachia így megmarad az allopatrikus fajok izoláló mechanizmusának.
Intermezzo. A kongresszus félidejében, a harmadik napon fél nap szünetet tartottunk. A szervezõk két fakultatív kirándulást kínáltak, de az Aarhusban maradóknak is bõven akadt látnivaló. Magam a Régi Városban töltöttem két felejthetetlen órát. Ebben az egyedülálló skanzenben egy komplett várost építettek fel a restaurált házakból, ahol a pékmûhelytõl a kalaposig, a postaállomástól a polgármester palotájáig és az egy tantermes, osztatlan iskoláig minden megtalálható úgy, ahogy az a XVIXIX. században volt.
A szünetet kihasználva (no meg esténként) a pár nap múlva kezdõdõ egyetemi félévre is készülõdni kellett már. A nagy kongresszusok augusztus végi idõpontja a lehetõségek által diktált hagyomány, hiszen a tanév alatt az egyetemi oktatók egy-két napnál hosszabb idõre nemigen utazhatnak el. A helyszín és a szervezõk viszont természetesen változnak: Aarhusban Volker Loeschcke vendégszeretetét élveztük, 1991-ben viszont az ESEB-sorozat emlékezetes harmadik kongresszusa Magyarországon volt, Vida Gábor elnökletével és Pásztor Erzsébet szervezésében.
Új utakon az életmenet-evolúció
Szembetûnõ volt az ez évi kongresszuson, hogy nem volt életmenet-evolúciós szekció (bár ilyen irányú elõadások szerepeltek más szekciókban). Furcsa, mert ezen a területen nagyon izgalmas eredmények vannak születõben, melyek akár az elmélet alapjait is forradalmasíthatják. Az életmenet-evolúció a szaporodással és demográfiával közvetlenül összefüggõ tulajdonságok, mint például a fészekalj- vagy alomméret, az utódgondozásba, illetve a szülõ saját fennmaradásába fektetett energia evolúcióját vizsgálja. Mivel ezek a tulajdonságok közvetlenül befolyásolják a rátermettséget, várható, hogy viszonylag erõs szelekció alatt állnak, és valóban a legrátermettebb változat az, amelyet a természetben megtalálunk.
A legrátermettebb, legjobb életmenetû változatok sajátságait évtizedek óta optimalizációsmodellekben vizsgálják. A legfrissebb empirikus eredmények és ezekbõl azért a kongresszuson is akadt néhány viszont arra mutatnak, hogy ez a klasszikus megközelítés nem mindig jó. Elõfordul, hogy nincs egyetlen legjobb változat; mert a különbözõ változatok rátermettsége attól függ, hogy jelenleg milyen egyedek, milyen gyakorisággal vannak jelen a populációban (gyakoriságfüggõ szelekció). Tapio Mappes (Finnország) és munkatársai például az erdei pockok vizsgálata során közvetlenül kimutatták, hogy a szaporodásba sok energiát fektetõ változat elõnyben van, ha a populáció többsége kevés energiát szán a szaporodásra, és fordítva, a kevés energiát befektetõk akkor vannak elõnyben, ha a többség sok energiát fektet be. Ez a fajta gyakoriságfüggés a két változat együttéléséhez vezet. Sajnos az még nem világos, hogy ebben az esetben pontosan milyen módon jut a ritka változat elõnyhöz. A tüskés pikó életmenetében U. Candolin (Svájc) és H.-R. Voigt (Finnország) rafináltabb gyakoriságfüggést talált. A kis testû hím pikók korábban foglalnak territóriumot a párzóhelyeken, míg a nagyobbak késõbb, a nõstényekkel együtt érkeznek. A korán érkezõk azonban a ragadozók miatt nagyobb veszélynek vannak kitéve. A kis hímek azért vállalják ezt a veszélyt, mert a nagyokkal egyszerre érkezve és velük versengve nem jutnának territóriumhoz és így szaporodni sem tudnának; elõbb érkezve viszont akadálytalanul elfoglalhatják a territóriumokat. A korai érkezés kockázatos stratégiájának természetesen csak akkor van értelme, ha nagy testû hímek elég nagy számban vannak a populációban.
Az ellenség belül van
Az életmenet-evolúció modelljei megmondják, hogy egy anyának optimálisan mennyi energiát kell az utódaira fordítania, ám az utódoknak errõl más véleményük van. Miért is ne szipolyozza ki egy utód a szülõjét, feltéve, hogy képes rá? A természetes szelekció önzõ logikája szerint az utód csakis azért érdekelt szülõje fennmaradásában, mert a szülõ újabb utódaiban részben a saját génjei lesznek jelen. A nemrég elhunyt William Hamilton egyik nagy felismerése volt, hogy egy génnek érdemes önzetlen viselkedésre késztetni az õt hordozó egyedet rokonaival szemben, akik legalábbis bizonyos valószínûséggel szintén hordozzák az illetõ gént. Az önzetlen viselkedés kiváltásával a gén saját kópiáit segíti másokban.
Igen ám, de az ördög a részletekben lakozik. Sok állatfajban az egy nõsténytõl születõ utódok más-más hímtõl származnak. Ilyenkor egy apai eredetû génnek nem érdemes a testvéreket segítõ önzetlen viselkedést kiváltania, hiszen a testvérek apja más, vagyis a testvérek nem hordozzák a szóban forgó gén kópiáit. Ugyanígy, az apai eredetû géneknek nem érdemes vigyázniuk az anyára, hiszen az anya jövendõ utódai már nem ettõl az apától kapnak majd géneket. Az anyai eredetû géneknek viszont érdemes segíteniük az anyai féltestvéreket és persze magát az anyát is.
Vajon tudhatja-e az utódban egy gén, hogy õ az apától vagy az anyától származik-e? A DNS-ben ez természetesen nincs kódolva, de a DNS-re utólag ráakasztott metilcsoportok mintázata eltérõ lehet a hímivarsejtekben és a petesejtekben. A különbözõ metilációs mintázatok más-más génexpresszióhoz vezetnek. A metiláció révén megvalósuló genetikai imprinting megjelöli az apai, illetve anyai eredetû géneket, és ez lehetõvé teszi, hogy a gének a származásuknak megfelelõen viselkedjenek.
David Haig (USA) Genetikai imprinting és a megosztott én címû élvezetes elõadásában amellett érvelt, hogy a genetikai imprinting egyenesen azért alakult ki, hogy a gének képesek legyenek differenciálni abban, hogy mely rokonokkal szemben gyakoroljanak önzetlenséget. Az önzetlenség fogalmát itt persze nagyon tágan kell érteni. Például az anyai eredetû gének eredetileg oda hatnak, hogy egy emlõsmagzat csak korlátozott mennyiségû tápanyagot vonjon el a méhlepényen keresztül (önzetlenség az anyával szemben). Az apai eredetû géneknek azonban nem érdekük az anyára vigyázni, ezek tehát növelik a tápanyagelvonást. Válaszul az anyai eredetû gének kevesebbet vonnak el (hiszen a tényleges elvonás az apai és anyai génhatások eredõje, s az anyai gének egy közepes szint megvalósítására törekednek), mire az apai gének persze még tovább fokozzák az elvonást.
Hogyan oldódik meg egy ilyen konfliktus? Az egyik kézenfekvõ lehetõség, hogy az anyai gének végül egyáltalán nem vonnak el tápanyagot. Ennél lejjebb azonban nem tudnak menni: ezután az apai gének szabadon növelhetik az elvonást, s így végül a hangosabban kiabáló gyõz.
Az apai és anyai eredetû gének ellentétes hatására közvetlen kísérleti adatok is vannak. Az újonnan megtermékenyített zigótában a sejtmagvak egyesülése elõtt kicserélhetõ egyik vagy másik sejtmag, s így olyan egérembriók is létrehozhatók, amelyeknek genetikailag két anyjuk vagy két apjuk van. Ezen embriók egyike sem életképes, de más-más okból. A két anyával (két petesejtmaggal) rendelkezõ utódoknál alig fejlõdik ki a méhlepény, így a magzatok éhen pusztulnak. Két apa esetén viszont az embrió alig alakul ki, csak a méhlepény növekszik. A különbözõ leszármazási vonalak keresztezésekor is tetten érhetjük az imprinting következményeit. Két, nagyjából azonos súlyú törzs keresztezésekor az utódok feltûnõen nagyok, ha az apa az elsõ, az anya a második vonalból származik, fordított keresztezésnél viszont az utódok szokatlanul kicsik. Ekkor arra gyanakodhatunk, hogy az elsõ vonal elõrébb tartott az apai és anyai gének közötti fegyverkezési versenyben. Az elsõ vonal apai eredetû génjei hangosabban kiabáltak tápanyagért, így a második vonal anyai génjei nem tudták kompenzálni õket; megfordítva, az elsõ vonal anyai génjei kiabálósabb apai génekre voltak felkészülve, mint amit a második vonalból kaptak.
Zárszavában David Haig a hölgyeknek kedveskedett: egyes jelek szerint az anyai eredetû gének növelnék a fej méretét a testhez viszonyítva (míg az apai gének hatása fordított lenne), sõt az agyvelõn belül az anyai gének növelnék a tudatos gondolkodást végzõ nagyagykéreg arányát, míg az apai gének a hipotalamusznak kedveznének egyelõre senki sem tudja, hogy miért.
Kommunikációs problémák
Az állati kommunikáció feltûnõ vonása, hogy az egymással közölt információ igaz, vagyis a kommunikáció becsületes. Természetesen nem arról van szó, hogy az állatok ne tudnák félrevezetni egymást, s ha a csalás elõnyös, akkor a természetes szelekció elterjeszti a csalókat. Ám végül mégiscsak a becsületes kommunikáció marad meg, mert az információt vevõ egyedek sem védtelenek: ha a hamis információból a vevõnek kára származik, akkor elõnyben vannak azok a vevõk, akik egyszerûen nem veszik figyelembe a kapott információt. A nem figyelõ vevõk elterjedésével a tulajdonképpeni kommunikáció megszûnik, s akkor már a csalóknak sem érdemes a közléssel strapálniuk magukat.
John McNamara (Nagy-Britannia) Alasdair Houstonnal közös elõadásában érdekes elemzéseket mutatott a kommunikáció evolúciójáról abban a kissé bonyolultabb esetben, ha az információt hordozó szignál zajos. A legmeglepõbb konklúziója azonban az volt, hogy még egy egyértelmû és becsületes információ is csökkentheti az információt kapó partner rátermettségét. Példaként egy egyszerû helyzet szolgál. A megtermékenyítés után a szülõk mindegyike eldöntheti, hogy gondozza-e az utódokat, vagy elhagyja õket. Ha mindkét szülõ az elhagyás mellett dönt, akkor az utódok elpusztulnak. Ha az egyik szülõ gondoz, a másik nem, akkor mondjuk három utódot lehet felnevelni. A befektetett munkának azonban ára van: a gondozó szülõ két (jövõbeli) utódnak megfelelõ fitnesst veszít. Végül ha mindketten gondoznak, akkor négy utódot tudnak felnevelni, de mindketten fizetik a gondozás árát. A megtermékenyítés után a nõsténynek világra kell hoznia az utódokat, s ez idõ alatt a hím már dönt, hogy marad-e vagy sem. A nõstény a hím döntésének biztos tudatában választhat, hogy mit csináljon. A hím el fogja hagyni az utódokat, a nõstény viszont gondozza õket. Ez a megoldás a nõstény számára egy utódnak megfelelõ fitnesst jelent (hármat nevel fel, de két egység árat fizet).
Mi lenne az eredmény akkor, ha az egyedek hasonló választás elõtt állnának, de semmit sem tudnának partnerük viselkedésérõl (még azt sem, hogy nõstény-e vagy hím)? Ekkor mátrixjátékkal állnánk szemben, amelyben az evolúciósan stabil stratégia 50 százalékos valószínûséggel dönt a gondozás mellett, s minden résztvevõ fitnesse 1,5 utódnak felel meg. A nõstény szempontjából ez jobb lenne, mint a becsületes információ mellett kapott 1 utód! Természetesen a nõstény itt nem sokat érne azzal, hogy nem veszi figyelembe a hím döntését. A helyzet kulcsa a hím kezében van, ha a hím már elment vagy hihetõen bejelentette, hogy elmegy , akkor a nõsténynek még mindig jobb, ha gondozza az utódokat.
A kommunikáció evolúciójával foglalkozó szekció utolsó elõadása igazi meglepetés. Rufus Johnstone (Nagy-Britannia) egy elmaradt elõadás helyett, beugróként beszél az extra információszerzés, hallgatózás evolúciójáról. Kiindulópontja az agresszivitás közismert játékelméleti modellje, a héjagalamb-játék. A játékban két ellenfél egy V értékû forrást (például táplálékot) szeretne megkapni. A héják teljes erõbedobással harcolnak, a galambok viszont az ellenfél agressziójának elsõ jelére átengedik a forrást. Egy héja és egy galamb találkozásakor tehát a héja lesz a nyertes. Két galamb békésen viselkedik egymással, egyikük (véletlenszerûen) megkapja a forrást, átlagosan V/2 nyereségük van. Két héja konfliktusában azonban véres küzdelemre kerül sor, egyikük megsebesül, ami miatt C-t veszít; átlagosan tehát csak V/2C/2 a nyereség. Ha C nagyobb, mint V, a héják és a galambok együtt élnek.
Johnstone modelljében az egyedek nem egyetlen héjagalamb-játékot játszanak, hanem sokat, mindig véletlenszerûen választott partnerral. A hagyományos héja- és galambstratégiák mellett bevezetünk egy hallgatózó (itt inkább megfigyelõ) stratégiát is, aki szemmel tartja a többieket. Ha mostani ellenfele az elõzõ fordulóban nyert, akkor a hallgatózó óvatosan galambként viselkedik; ha viszont az ellenfél az elõbb veszített, akkor a hallgatózó héjaként támad rá.
Azt gondolhatnánk, hogy a hallgatózó stratégia mérlegelésével csökkenti a véres küzdelmek számát, de ez nincs így. A három stratégia egyensúlyában a véres küzdelmek száma nagyobb, mint a klasszikus héjagalamb-egyensúlyban! A magyarázat utólag pofonegyszerû: a hallgatózók miatt az egyedek most már nemcsak a forrásért küzdenek, hanem a reputációjukért is. Ha most nyerek, a forrás is az enyém, és ráadásul a hallgatózó is szelíden fog viselkedni velem a következõ fordulóban!
Finálé. Az Evolúcióbiológiai
Társaság esti közgyûlésén csupa
jó hír hangzott el. Jacqui Shykoff titkár a
taglétszám jelentõs gyarapodásáról
számolt be (tündéri hét hónapos kisbabájával
az ölében), s a Társaság által jegyzett
szakfolyóirat (Journal of Evolutionary Biology) példányszáma,
terjedelme és idézettsége is szépen növekszik.
A kongresszussorozat is nagyon népszerû, olyannyira, hogy
idén a magas túljelentkezés miatt a kései regisztrációkat
már nem is fogadták el, az elõadótermek így
is zsúfolásig megteltek. A búcsúzás
körül a hogyan továbbról, a kongresszus jövõbeli
szervezési lehetõségeirõl beszélgettünk.
Egy biztos: viszontlátásra két év múlva,
Angliában.
IRODALOM
A kongresszus programja és az elhangzott elõadások kivonatai megtalálhatók a https://www.biology.au.dk/eseb/program.html weboldalon. Cikkemben a következõ elõadásokat használtam fel:
Roger K Butlin, Craig Wilding & John W Grahame: Differential gene exchange between morphs of Littorina saxatilis detected using AFLP markers (9. szimpózium)
U. Candolin & H.-R. Voigt: Selection on arrival date at breeding areas in sticklebacks: Why do small males arrive first? (7. szimpózium)
Andrew G. Clark: Population genomics (plenáris)
Martijn Egas & Filipa Vala: Wolbachia and speciation (9. szimpózium)
David Haig: Genomic imprinting and the divided self (plenáris)
D. C. Jeffares, A. M. Poole, D. Penny & P. Arctander: The origin of messenger RNA in the RNA world: Introns first, exons later (3. szimpózium)
Rufus Johnstone: Eavesdropping and animal conflict (11. szimpózium)
Tapio Mappes, Minna Koivula, Esa Koskela, Tuula A. Oksanen & Barry Sinervo: Negative frequency dependent selection on reproductive effort (7. szimpózium)
John M. McNamara & A. Houston: The role of information in evolutionary game theory (11. szimpózium)
Andrew Read: The evolution of malaria (plenáris)
| Természet Világa, | 132. évfolyam, 12. szám, 2001.
december
https://www.chemonet.hu/TermVil/ https://www.kfki.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez