Szállítószalag vagy szerelőcsarnok?
A Golgi-készülék működésének új modellje
A Golgi-készülék a sejt által készített fehérjéken végzi el az utolsó simításokat, válogatja, csomagolja őket. Eddig azt gondolták, hogy úgy működik, mint egy szállítószalag: sejtmag felőli oldalán bekerülnek a fehérjék, ciszternáiban tovaszállítódnak, külső oldalán pedig becsomagoltan távoznak. A legújabb kutatási eredmények alapján a Golgi-komplex saját fehérjéi kis hólyagocskákban visszaforgatódnak, miközben a szállítandó fehérjék látszólag helyben maradnak az érő ciszternákban. Így az új nézet szerint a Golgi-készülék inkább egy olyan szerelőcsarnokhoz hasonlít, ahol a munkadarab egy helyben áll, míg az újabb munkafázisokat végző robotok cserélődnek körülötte. A Golgi vázfehérjéi sem képeznek statikus, merev hálózatot, mert a citoplazmán keresztül dinamikusan visszaépülnek a Golgi-apparátusba. Ez az újabb modell talán közelebb visz bennünket a Golgi-készülék igazi szerkezetéhez, és érthetőbbé teszi működését.
Camillo Golgi (1843–1926), miközben a róla elnevezett fémimpregnációs technikával idegsejteket tanulmányozott, kosárszerű hálózatot figyelt meg a sejtek magja körül (1. ábra). 1898-ban tette közzé az első leírást erről a szerkezetről, melyet „belső hálózatos készüléknek” nevezett el (apparato reticolare interno). Ma Golgi-készüléknek, Golgi-apprátusnak, Golgi-komplexnek, vagy egyszerűen csak Golginak hívjuk e sejtszervecskéket.
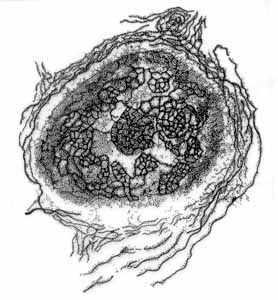
1. ábra. Golgi rajza az általa felfedezett "sejten belüli hálózatos készülékről" (apparato reticolare interno), vagyis a Golgi-apparátusról, ahogy egy ló csigolyaközi dúcából származó idegsejtben látta
A Golgi-készülék lapos, szorosan egymás mellé rendezett ciszternák csoportjából, az úgynevezett diktioszómákból áll. Egy-egy csoportban általában 5-10 párhuzamos ciszterna figyelhető meg. Ezek membránjain nincsenek riboszómák, ezért a készülék fehérjét nem szintetizál. Egy-egy ciszternacsoporton belül az egyes ciszternákat határoló membránok vastagsága jellegzetesen változik: a sejtmag felőli szélső ciszterna esetében ez körülbelül az endoplazmatikus retikulum (ER) membránvastagságával megegyező, azaz 6 nm, míg a másik oldalon 10-12 nm, ami a plazmamembránhoz hasonló. Az előbbit cisz vagy bemenő oldalnak, míg a másikat transz vagy kimenő oldalnak nevezik. A két szélső ciszterna között elhelyezkedő mediális ciszternák határoló membránjainak vastagsága cisz–transz irányban fokozatosan nő. Az egyes ciszternák szélein, különösen a transz oldalon, kisebb-nagyobb tágulatok vannak: ezek váladékkal teli vakuólumok, melyek aztán lefűződnek.
Komplex kérdések
A Golgi-készülék történetének meghökkentő vonása, hogy tele van ellentmondásokkal. Először is, ötven évig kemény vita folyt arról, hogy a Golgi valódi sejtszervecske-e vagy csak műtermék. A kétség azért merült fel, mert a Golgi-apparátus élő sejtekben fénymikroszkóppal nem volt látható, csak Golgi „fekete reakciónak” (la reazione nera) nevezett nehézfémfestéses eljárásával lehetett megfigyelni. Ezt azonban nehéz volt megbízhatóan reprodukálni és sok más struktúrát, például egész idegsejteket is megfestett. Abban az időben, mikor Golgi felfedezte a Golgi-készüléket, maga is vitában állt Ramón y Cajallal (ejtsd: ramon i kahal), a híres spanyol neuroanatómussal abban a kérdésben, hogy a neuronok folytonos vagy megszakított hálózatot képeznek. A vitát ugyan elvesztette, de 1906-ban osztozott Cajallal az élettani és orvosi Nobel-díjon. Az elismerést nem a Golgi-készülék felfedezéséért kapta, hanem a „fekete reakció” bevezetéséért az idegszövet tanulmányozásába. Ezt használta Cajal is, hogy bebizonyítsa, nincs igaza Golginak, mert az idegsejtek különálló egységek. Érdekes, hogy mind a mai napig a Golgi-festés módosított változatát használják egyes idegsejtek és neuronhálózatok megfestésére a neurobiológusok.
Mikor szűnt meg a Golgi-készülék puszta létezése körüli kétely? Akkor, amikor bevezették az elektronmikroszkópos vizsgálatokat a sejt szerkezetének tanulmányozására. A Golgi-készülék az elektronmikroszkópos képen görbült, sima felszínű ciszternák halmazaként tűnt elő, amelyet különböző méretű vakuólumok vettek körül. Hasonló megfigyelések egész garmadája született az 1950-es években, melyek bizonyították, hogy a Golgi-készülék a sejtekben általánosan jelen van, és megmutatták belső változatosságát és összetettségét. Ettől kezdve a Golgi-készülék az érdeklődés középpontjába került a sejtbiológiában. Következzen most néhány mérföldkő a Golgi-kutatásban és néhány újabb ellentmondás, amely a komplex tanulmányozása közben felvetődött.
Az 1960-as évekig sok feltételezés, de kevés tényleges információ állt rendelkezésre a Golgi-apparátus feladatáról. Már régen észrevették a kutatók, hogy a Golgi erősen fejlett a mirigysejtekben, de egészen a hatvanas évekig nem vált világossá a szekrécióban és glükozilálásban betöltött szerepe. Új technikák, mint a sejtfrakcionálás és az elektronmikroszkópos autoradiográfia bevezetése tette lehetővé ezek felismerését. E két módszert alkalmazta Palade a hasnyálmirigy külső elválasztású sejtjein, és egymást kiegészítő biokémiai és morfológiai adatokat kapott a Golgi részvételéről a szekréciós fehérjék irányított transzportjában.
A hatvanas évek egy másik alapvető felfedezése az volt, hogy rájöttek, a Golgi-készülék jól körülírható alrendszerekből, szubkompartmentumokból épül fel. Ezt az ólom-foszfát alapú enzimcitokémiai módszerek elektronmikroszkópos alkalmazása tette lehetővé. Novikoff és Goldfisher vették észre először, hogy a savas foszfatáz és tiamin pirofoszfatáz enzimek nem egyenletesen oszlanak el a Golgi-komplexben, hanem a kimenő (transz) oldalon koncentrálódnak. Hasonlóan más foszfatázoknak is megkülönböztetett helye van a komplexen belül.
Mit csinál a Golgi-készülék?
Mivel a Golgi-ciszternák és az endoplazmatikus retikulum üregrendszere nincs közvetlen kapcsolatban egymással, ezért a retikulum lumenében megjelenő anyagok nem tudnak közvetlenül átömleni a Golgi ciszternarendszerébe. A két kompartmentum között az anyagáramlás vezikuláris transzport segítségével valósul meg (2. ábra). Ennek során az endoplazmatikus retikulum Golgi-készülékkel határos membránjain apró kitüremkedések képződnek, melyekről 50-80 nm átmérőjű vezikulák fűződnek le. Ezek a Golgi cisz oldalához sodródnak és ott egybeolvadnak az első ciszternával. A folyamat kezdő lépését, a membránkitüremkedések megjelenését különleges burokfehérjék, a COP-fehérjék (coat protein, „kabát” fehérje) indítják be. (A sejtbiológiában szokásos angol rövidítésekhez híven itt is egy szójátékkal találkozunk: cop angolul zsarut, illetve letartóztatást jelent.) Ezek a fehérjék nemcsak a vezikulaképződést indukálják, hanem védik is a vezikulát a véletlenszerű visszaolvadástól. A vezikula membránja tartalmaz egy azonosító fehérjét is, mely nagy affinitással képes a cisz-Golgi-membránokon levő jelfogó fehérjéhez kötődni. Ez a jelfogó rendszer biztosítja azt, hogy a vezikulumok a donor kompartmentumról leválva csakis a számukra specifikus fogadó (akceptor) kompartmentum membránjával tudjanak fuzionálni. A jel–jelfogó kapcsolat kialakulása mellett a membránösszeolvadás kiváltásához még egy fehérjecsoport jelenléte szükséges (bővebben lásd folyóiratunk 2001. novemberi számában). Vezikuláris transzport nemcsak az endoplazmatikus retikulum és a Golgi-komplex között működik; vezikulák szállítják az anyagot a transz-Golgi-hálózatból az endoszóma/lizoszóma-rendszerbe és a sejtfelszínről az endoszómába is. Kimutatható, hogy vezikuláris transzport visszafelé, a Golgi-endoplazmatikus rendszer irányba is működik: a burkos vezikulák egy csoportja, (a COP I burokkal borított változat) képes a Golgi-ciszternákból az endoplazmatikus retikulum saját fehérjéit visszaszállítani (2. ábra).
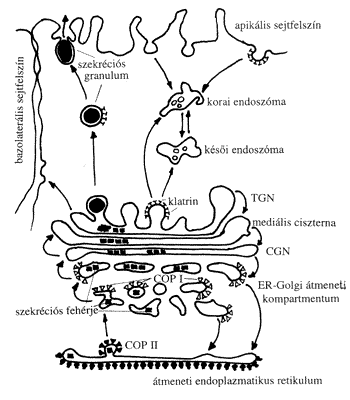
2. ábra. Főbb szállítási útvonalak a Golgi-készülékben (CGN – cisz-Golgi-hálózat,
TGN – transz-Golgi-hálózat, ER – endoplazmatikus retikulum, COP I,II –
I-es, II-es típusú burokfehérje, a további magyarázatot lásd a szövegben)
A vezikuláris forgalom a Golgi-készülékben két fő útvonalhoz csatlakozik: az exocitózis és az endocitózis irányába (2. ábra). A váladéktermelő sejtekben a kiválasztandó fehérjék, a membrán-glükoproteidek és a lizoszomális enzimek poliriboszómákon képződnek és először az ER-be kerülnek, ahol poszttranszlációs módosításokon esnek át. Az ER-ből membránjukon különleges burokfehérjéket, COP II fehérjéket viselő hólyagocskákba csomagolva lépnek ki. Ezek arra szolgálnak, hogy a fehérjéket az ER – Golgi-komplex átmeneti kompartmentumba vigyék. Innen a régebbi megfigyelések szerint COP I fehérjékkel burkolt vezikulák szállítják őket a cisz-Golgi-hálózatba (cis-Golgi network, CGN). Visszafelé irányuló membránszállítást (retrográd transzport) is megfigyeltek a felsőbb Golgi-ciszternák között. A fehérjék különböző szekréciós utakra történő válogatása a transz-Golgi-hálózatban (trans-Golgi network, TGN) történik. A lizoszomális enzimek például mannóz 6-foszfát receptorokhoz kötődnek a Golgiban, klatrin burkolatú hólyagocskákba csomagolódnak a transz-Golgi-hálózatban és a korai vagy késői endoszomákhoz szállítódnak. A szekréciós- és membránfehérjék szintén a transz-Golgi-hálózatban válogatódnak ki és szekréciós granulumok formájában jutnak el a csúcsi (apikális) sejtfelszínhez. A polarizált mirigysejtekben a hólyagocskák elkülönült útvonalon szállítódnak az alapi és oldalsó (bazolaterális) sejthártyaszakaszokhoz (2. ábra).
Működési modellek
A hatvanas, hetvenes évek óta világosan látjuk, hogy a szállítandó fehérjék keresztüljutnak a Golgi-apparátuson, de negyven éve vita tárgya, hogy ez a folyamat milyen mechanizmussal valósul meg. Két modell alakult ki. Az egyik az érési vagy ciszternaprogressziós modell, mely Morrétől és Leblandtól származik az ötvenes évekből. Ez a Golgi-készüléket egy palackozóüzemhez hasonlítja: a ciszternák kialakulnak a bemenő vagy cisz oldalon, és fokozatosan mozognak előre a kimenő vagy transz oldal felé, ahol a csomagolásban felhasználódnak.
A hetvenes évek új felfedezéseit, a szubkompartmentumokat és a membrán-visszaforgatás jelenségét nehéz volt a érési modellel magyarázni. A vezikuláris transzport/állandó ciszterna modellt már ezen új információk figyelembevételével alakították ki 1981-ben. Palade és munkatársainak kutatásai óta az ER-ből a Golgi-készüléken keresztül megvalósuló fehérjeszállítást olyan folyamatnak tekintették, melyet kis szállító vezikulák hajtanak végre. Eszerint minden ciszterna vagy ciszternacsoport elkülönült összetételű kompartmentet alkot, melyek között az anyagforgalom vezikulák segítségével történik. Ezek leválnak az egyik kompartmentről és egybeolvadnak a másikkal. Ez a modell in vitro rendszerekben jól működött, így tizenöt évig uralta a területet. Az utóbbi években, a legújabb eredmények alapján azonban komoly kihívásokkal kell szembenéznie. Egyrészt nem sikerült szekréciós fehérjét találni a szállító hólyagocskákban. A modell a prokollagén szekréciójának vizsgálatával is megkérdőjeleződött. Ez a molekula egyszerűen túl nagy (1500 aminosav) ahhoz, hogy egy átlagos 60-70 nm átmérőjű szállító vezikulába beleférjen. Nem könnyű ezzel a modellel a ciszternákban található összetett struktúrák (pl. kazein szubmicellák, algapikkelyek, virális membrán glikoproteinek) érését sem megmagyarázni, ezért felül kellett vizsgálni a meglévő elképzeléseket.
A ma elfogadott változat az eddigiek hibridje. Az előre irányuló szállítás a ciszternák érésével történik, ehhez kapcsolódik a Golgi-enzimek és membránok visszaforgatása vezikulák segítségével. A ciszternák az endoplazmatikus retikulából származó vezikulák, illetve tubuláris intermedierek összeolvadásával keletkeznek, és cisz-transz irányban érnek azzal, hogy az ellentétes irányban mozgó szállító hólyagocskák segítségével kicserélődnek bennük az enzimek. Azóta ez az elmélet egy sokkal kisebb molekulára, a vezikuláris sztomatitisz vírus egyik transzmembrán fehérjéjére is igazolódott. Azt találták, hogy a szállítandó molekulák közül egy sem kerül bele a vezikulákba, amíg a Golgi-készüléken áthalad és mindegyik azonos ütemben szállítódik, azaz egy-egy érő ciszternában foglal helyet. Az elmélet más oldalait biokémiai és morfológiai bizonyítékok támasztják alá. A legfontosabb, hogy a Golgi-készülék saját fehérjéi visszaforgató vezikulákba rendeződnek a ciszternahalom különböző szintjein (2. ábra). A COP I-fehérjét tartalmazó hólyagocskákban vagy a cisz-, vagy a mediális-Golgira jellemző jelölő fehérjék vannak, de alig található bennük érő szekréciós fehérje. Ezek a vezikulumok a szállítandó fehérjék kiválogatásához a GTP hidrolízisével nyerik az energiát.
Golgi-mátrix
Az eredmények, melyek szerint a legtöbb, de talán az összes szállítandó fehérje az érő ciszternákban halad előre, míg az enzimek a COP I burkolatú vezikulákban vannak jelen, alátámasztják a Golgi-transzport érési modelljét. Ez persze újabb kérdéseket vet fel a Golgi-készülék egymásra rétegzett ciszternákból álló szerkezetének fenntartásával kapcsolatban. Az érési modell azt feltételezi, hogy a szerkezeti fehérjekomplexek, melyeket gyakran Golgi-mátrixként emlegetnek, szintén dinamikusan visszaforgathatók a ciszternák érése közben. Korábban a hosszú életű ciszternák viszonylag stabil alkotóelemének tartották őket. A feltételezett Golgi-mátrixot eredetileg olyan fehérjehálózatként írták le, mely izolálva a Golgi-membránokról megtartja a ciszternás szerkezetét és specifikusan képes Golgi-enzimeket kötni. A tisztított Golgi-membránok valószínűleg azért őrzik meg jellegzetes, egymásra rétegzett ciszternás szerkezetüket, mert a mátrixfehérjék összefüggő vázat alkotnak, mely megtartja ezt az egyedi szervezettséget (3. ábra).
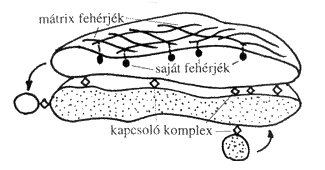
3. ábra. A Golgi-mátrix szerkezete
A Golgi-mátrix több tagja csavart spirált tartalmazó fehérje (erről a molekuláris szerkezetről folyóiratunk 2001. novemberi számában olvashattak). Ezek a membránban ülő fehérjék szabad végükkel szorosan egymáshoz kapcsolódhatnak és így membránokat, sőt sejtszervecskéket is képesek egymáshoz húzni. Régebben azt feltételezték, hogy a Golgi-mátrix szerkezeti fehérjék stabil együtteséből áll, és részt vesz a Golgi-membránok szerveződésének és azonosságának kialakításában. Nyilvánvaló, hogy csak a mátrix nem elegendő a Golgi-készülék megszervezéséhez. Ha például gátoljuk a Golgi-ciszternák utánpótlását (a COP-II fehérjével burkolt vezikulumok útvonalának blokkolásával, 2. ábra), egy idő után az egész Golgi-készülék eltűnik, visszaforgatódik az endoplazmatikus retikulumba. Ez természetesen a teljes fehérjeszállítást is megbénítja a sejtben, hiszen nincs működő Golgi, ami ezt elvégezné. Bár a Golgi-mátrix nagyon fontos a komplex szerkezetének és az egyes kompartmentek azonosságának fenntartásában, mégsem bizonyított egy statikus, térben is rendezett szerkezetű, valóban „mátrix-szerű” anyag jelenléte. Inkább egy meglehetősen dinamikus modellt kell elképzelnünk, melyben a szerkezeti fehérjék kisebb komplexekbe verődve jelölik meg a szubkompartmenteket (3. ábra).
További talányok
Mivel a szekréciós útvonal elsősorban teherszállító rendszer, kézenfekvőnek látszik enzimek kis adagjait a nagy mennyiségű módosítandó szubsztráthoz vinni. Sokkal hatékonyabbnak tűnik ez a megoldás, mint az ellenkezője, vagyis nagy mennyiségű teher szállítása kis vezikulákban. Mindazonáltal még mindig sok a nyitott kérdés, melyeket ez a modell sem old meg. A Golgi-enzimek visszakerülnek az endoplazmatikus retikulába, miközben minden előre haladó szállítmány (ide értve az enzimeket is) azonos ütemben mozog a Golgi-komplexben előre. Ha valóban így áll a helyzet, akkor a cisz-Golgi-ciszternák nemcsak cisz-enzimeket, hanem jelentős mennyiségű visszaforgatott mediális- és transz-Golgi-enzimet is tartalmaznának, melyek a nekik megfelelő helyre igyekeznek. Hogyan magyarázható mégis, hogy ezek az enzimek nem egyenletesen oszlanak el a Golgi-komplexben? A legegyszerűbb magyarázat az volna, hogy lépcsőzetes enzim-újrahasznosítás működik, mely egyszerre csak a következő ciszternáig viszi a vezikulákban az enzimeket ahelyett, hogy egyenesen visszaengedné őket az endoplazmatikus retikulumba. Az tenne lehetővé egy ilyen mechanizmust, ha e vezikulák nem mozoghatnának szabadon, hanem kialakulásuktól kezdve a szomszédos célkompartmenthez lennének kikötve. Így azonban csak elodáztuk a probléma megoldását, mivel a különböző Golgi-kompartmentek részletes molekuláris térképe még nem készült el.
Egy másik kérdéskör a feltételezett Golgi-mátrix fehérjéinek specifikus célbajutását érinti, valamint azt, hogyan tudják megőrizni helyüket a különböző ciszternákban. Egyes mátrixfehérjék a citoplazmán keresztül juthatnak vissza, de az integráns membránfehérjék csak vezikulákban forgathatók vissza.
A szemléletváltás ellenére, mely a Golgi-készüléket ma már nem statikus
sejtszervecskének, hanem dinamikus szerkezetnek tekinti, az alapvető sejtbiológiai
kérdés megmarad: hogyan tartható fenn ez a struktúra az endoplazmatikus
retikulumtól a sejtfelszínig tartó nagy szekréciós anyagáramlásban? A Golgi-készülék
történetéből megtanulható az, hogy új megközelítésre és különféle területekről
származó összefüggő információkra van szükség ahhoz, hogy általánosan elfogadott,
pontos modellt alkothassunk.
Irodalom
1. F. A. Barr (2002) The Golgi apparatus: going round in circles? Trends
in Cell Biology 12: 101–104.
2. M. G. Farquhar és G. E. Palade (1998) The Golgi apparatus: 100 years
of progress and controversy. Trends in Cell Biology 8: 2–10.
| Természet Világa, | 133. évfolyam, 10. szám, 2002. október
https://www.chemonet.hu/TermVil/ https://www.kfki.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez