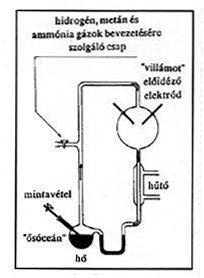
különbözô egyszerûbb szerves
molekulák prebiotikus
szintézisére alkalmas
| EVOLÚCIÓ |
CSERMELY PÉTER
Az élet születésének
biokémiája
Elképzelések
arról, hogyan vezethetett
a molekulák evolúciója az élet keletkezéséhez
Bizonyos becslések szerint az élet kb. 3,5–4 milliárd évvel ezelôtt alakult ki a Földön. Mi történt azalatt a kb. félmilliárd év alatt, amely a földkéreg megszilárdulásától az elsô primitív egysejtûek megjelenéséig telt el? Írásomban az élet keletkezésének megfejtésére irányuló "nyomozómunka" néhány frissebb eredményét foglalom össze, rámutatva a legfontosabb nyitott kérdésekre és nem elhallgatva saját véleményemet, spekulációimat sem.
A Föld a szervetlen molekulák idején
Az élet születésének kb. félmilliárd éve [1], az ún. "prebiotikus evolúció" során nem az élôlények harcoltak a fennmaradásért folyó küzdelemben, hanem kémiai, biokémiai reakciók serege zajlott egymással párhuzamosan az adott földi körülményeknek legmegfelelôbb makromolekulák és reakcióutak fennmaradását eredményezve. A "reakciók csatájának" kezdetén a Földet túlnyomó részben szervetlen molekulák uralták. A légkörbôl hiányzott az oxigén, a mainál sokkal redukálóbb atmoszféra nitrogént, vízgôzt, szén-dioxidot, szén-monoxidot, hidrogént, esetleg ammóniát és metánt tartalmazhatott. Mivel a szén-dioxid mennyiségét, és ezáltal a földi ôslégkört felmelegítô üvegházhatást éppúgy nehéz megbecsülni, mint a vulkáni tevékenység kiterjedését, nincsenek pontos fogalmaink arról, hogy a földi ôsóceán mennyiben volt folyékony, illetve fagyott. A földköpeny számos olyan szilikátból, agyagásványból, piritbôl és más anyagokból álló felületet kínált, amely alkalmas volt bizonyos kémiai reakciók szelektív meggyorsítására, katalízisére. Ezek voltak a legfontosabb elemei annak a színtérnek, amelyen az élet keletkezéséhez vezetô folyamatok zajlottak. Mielôtt azonban az élethez vezetô út lehetséges lépéseit sorra venném, két megjegyzést szeretnék tenni.
Az elsô megjegyzés a megismerés korlátait szeretné felvázolni. Az elôbbiekbôl is látszik, hogy a prebiotikus evolúció kutatói már a kezdet kezdetén is számos, szinte áthághatatlan nehézségbe ütköznek. Egyrészt az eltelt évmilliárdok a korai történések szinte minden nyomát elmosták. Másrészt a prebiotikus evolúciót jellemzô folyamatok rekonstrukciójához, modellezéséhez legalább két szemléleti, megközelítésbeli korlátot is le kell dönteni: az egyik az idôkorlát. Az a kémiai reakció, amely kb. százezer évet igényel ahhoz, hogy közel teljesen végbemenjen, mai pergô agyunknak reménytelenül lassú. Ugyanakkor ez a százezer év az élet kialakulására rendelkezésre álló idônek mintegy ötezred része (az élet teremtésének napjából mintegy 17 másodpercet foglal el). Az idôkorlát a folyamatok befejezettségében is jelentkezik. A mai élôvilágot szemlélve óhatatlanul annak viszonyait, megoldásait akarjuk visszavetíteni a teljesen más körülmények között kialakult régmúltba. Nagyon nehéz, emberpróbáló szemléleti kérdés, hogy a mai aggyal szinte rekonstruálhatatlan, sok száz millió évvel ezelôtt nyomtalanul eltûnt, primitív formációkat az akkori fejlettségre és viszonyokra helyes választ adó, szinte egyeduralkodó megoldásokként fogjuk fel.
A másik korlát térbeli. Az emberi gondolkodás hüvelykekben, lábakban, méterekben mért dimenzióihoz már az egész Földre kiterjedô egynemû, homogén modell alkotása is nehéznek bizonyulhat. Az élet keletkezésében azonban az irdatlan bolygóméreteknek olyan kicsi, helyi elemei játszhatták a legnagyobb szerepet, mint a megbecsülheteten számú vulkánok óceánmosta lejtôi, vagy a nem tudjuk hol és nem tudjuk mikor éppen kiszáradófélben lévô lagúnák. Így az élet kleetkezésének magyarázatában az egész Földre kiterjedô homogén modellnek értelme sincs.
A második megjegyzés az egész eszmefuttatás esetleges haszontalanságáról szól. Arrhenius és Hoyle nyomán legismertebben a Nobel-díjas Francis Crick fogalmazta meg az ún. pánspermia elméletet [2], amely leegyszerûsítve azt feltételezi, hogy az élet egy meteorit felszínén került a Földre és ott kedvezô körülményeket találva elszaporodott. Az a nemrégi felfedezés, amely során az Antarktiszon 1984-ben megtalált, 13 000 évvel ezelôtt a Földbe csapódott, marsi eredetû ALH 84001-es meteorit belsejében az ôsi, marsi élet egysejtûinek nyomaira bukkantak [3], az elsô konkrétabb bizonyítékát adta annak, hogy idegen eredetû élôlények a Földre kerülhettek. Bár az adott meteoritban rejtôzô baktériummaradványok már régesrég elhaltak, mire a meteor a Földet elérte és mire a meteor ideért, a földi élet mai formája már meglehetôsen jól kialakult. Ennek ellenáre nem kizárt, hogy más meteorokon utazó, "szerencsésebb" társaik benépesíthették ôsi bolygónkat. A pánspermia elmélet az élet keletkezésének kérdését persze nem válaszolja meg, csak a labdát dobja egy bolygóval (naprendszerrel) odébb. Ebbôl a szempontból az elôzô bekezdésben említett bizonytalanságok még szerencsének is tekinthetôk, hiszen az ekkora elhanyagolásokkal született elméletek nem csak a Föld, hanem számos hasonló bolygó felszínén megjelenô élet magyarázatául is szolgálhatnak. Jelenlegi tudásunkkal megbecsülhetetlen annak a valószínûsége, hogy volt-e, voltak-e olyan alkalmak, amikor a nem földi eredetû élet nyomai a Földre kerültek. Ha volt ilyen esemény, a legvalószínûbb feltételezés az, hogy az újonnan érkezett formáció bekerült az addigi reakciók, életformák versenyébe, ahol vagy nyert és átalakította a fejlôdés menetét, vagy vesztett és nyomtalanul, vagy bizonyos kisebb változások nyomát maga után hagyva eltûnt.
Mi kell ahhoz, hogy egy szervezet élô legyen? A legprimitívebb élô szervezetnek is rendelkeznie kellett egy olyan örökítô anyaggal, amely a szervezet mûködéséhez szükséges információt az utódokra átörökítheti. A környezete változásaitól való függetlenedéséhez szüksége volt egy határoló hártyára, membránra. Végezetül a membránnak az osztódással párhuzamos növekedéséhez és az örökítô anyag kettôzôdéséhez már az elsô élô szervezeteknek is ki kellett fejlesztenie egy olyan primitív anyagcsererendszert, amely saját magát szabályozni képes (lásd Manfred Eigen, illetve Gánti Tibor és Szathmáry Eörs munkáit [4–6]). Azt az egyszerûsítést is meg lehet kockáztatni, hogy molekuláris szinten az élethez kell valami, amelyrôl közel hibamentesen másolatok készíthetôk (ezt a továbbiakban templátnak fogom nevezni) és kell valami, amely a kémiai folyamatokat felgyorsítani, szabályozni, irányítani képes (a továbbiakban: katalizátor).
Elméletek az egyszerû szerves molekulák keletkezésére
|
| 1. ábra. A Miller-Urey reaktor különbözô egyszerûbb szerves molekulák prebiotikus szintézisére alkalmas |
Olyan szerves molekuláknak, mint a fehérjealkotó aminosavak közül a glicin, az alanin, az aszparaginsav és a glutaminsav, illetve az anyagcsere-folyamatokban részt vevô tejsav és borostyánkôsav az ôsi légkörhöz hasonló körülmények közötti gyors és hatékony szintézisét elôször Harold Urey tanácsait felhasználva Stanley L. Miller valósította meg. Miller az ôsatmoszférának megfelelô gázelegyben a villámlásoknak megfelelô elektromos kisüléseket hozott létre (1. ábra [7]). Úttörô kísérleteit számos hasonló próbálkozás követte, ahol a kiindulási gáz-, illetve folyadékelegy összetételét, a közölt energia jellegét (elektromágneses sugarak, illetve a közel 4 milliárd évvel ezelôtti Földet ért gyakori meteorbecsapódások [8]) és a jelenlévô katalizátorokat variálták. Így az aminosavak és a kis szénatomszámú zsírsavak mellett a nukleotidbázisok (pl. az adenin) szervetlen molekulákból, pl. hidrogén-cianidból való keletkezését is igazolni lehetett [5,9,10]. Formaldehidbôl egyszerû reakciók segítségével cukrok egész serege, így pl. ribóz állítható elô [9–11]. Ha a fentiekhez hozzávesszük a legkülönbözôbb molekulák felület-(pirit)-katalizált keletkezésének Wächtershäuser által javasolt útját [11], akkor az ôsi földi légkör elemi alkotóiból szinte minden olyan egyszerûbb szerves molekulát elô lehet állítani, amely egy képzelt primitív lény életfolyamataihoz nélkülözhetetlen. Bizonyos szerves molekulákat a Földünk légterébe lépô üstökösök is magukkal hozhattak [8]. Az építôkövek megjelenése után a következô lépés valószínûleg az információhordozó makromolekulák kialakulása volt.
Fehérje és ribonukleinsav: a tyúk–tojás probléma
Az élet kialakulásához az elôzôek szerint molekuláris szinten egy templát és egy katalizátor jelenléte szükséges. A kémiai reakciókat meggyorsító és szelektivitásukat biztosító fehérjéket hosszú ideig az élet egyik legfontosabb alkotóelemének, feltételének tekintették. E kép kialakulásához a század hatvanas és hetvenes éveiben a fehérjebiokémia diadalmenetén kívül az is hozzájárult, hogy különösen Sidney Fox munkássága nyomán [12] a fehérjék aminosavakból történô szintézisének meglehetôsen egyszerû módjait sikerült felfedezni. Nem volt olyan adat azonban, amely fehérjérôl fehérjét másoló rendszer létére engedett volna következtetni. Ismertük a katalizátort, de hiányzott a templát. A hetvenes évek végétôl az élettudományokra a nukleinsavakkal foglalkozó molekuláris biológia térhódítása nyomta rá a bélyegét. A nukleinsavak, különösen az ôsi körülményeknek jobban megfelelô ribonukleinsav (RNS) ideálisak a templát szerepére. A fehérjék "trónfosztása" a ribonukleinsavak katalitikus szerepének felfedezésével [13] következett be. A korábbi, "Hogyan alakították ki a nukleinsavakat a létezô primitív enzimek?" kérdés helyébe a "Hogyan szintetizálódtak a fehérjék a primitív, csak ribonukleinsavakat tartalmazó riboszómákon?" felvetés lépett. A ribóz viszonylagos labilitása [1] azonban a tisztán ribonukleinsav-alapú földi ôsvilág létét meglehetôsen megkérdôjelezi. A fehérjéknek a nukleinsavakhoz hasonlatos szabdalódását (splicing mechanizmusát) már 1990-ben felfedezték [14]. Ez az átalakulás bizonyos fehérje-templát szerepre enged következtetni. Leenek és munkatársainak az a nemrégi felfedezése, amely arról számolt be, hogy a kis fehérje peptidek képesek elôsegíteni önmaguk kialakulását [15], újabb példát adott arra, hogy primitív körülmények között fehérjék igenis mûködhettek templátként. A fehérje–RNS mérkôzésben a fehérjék tehát kezdenek az egyenlítés felé haladni.
A fehérjék, enzimek kialakulása
Matthews és Moser harminc évvel ezelôtt
hidrogén-cianidból, ammóniából és
vízbôl kisebb pepideket állított elô [16].
Aminosavak polimerizációjával Akabori és
Oró is képes volt közepes molekulatömegû
(kb. 2 kDa-os), fôleg glicint tartalmazó polipeptideket szintetizálni
[5,9]. A fehérjetermészetû anyagok közvetlen elôállításában
azonban Sydney Foxnak már a bevezetôben is említett
kísérletei jelentették a legnagyobb elôrehaladást.
Fox a természetes aminosavak glutaminsavban és aszparaginsavban
feldúsított keverékét 160–200 fokon néhány
órán át hevítette. A keletkezett olvadékot
vízben oldva akár 20 kDa molekulatömegû ún.
"proteinoid"-ot kapott [12]. A különleges elnevezés
arra utal, hogy a képzôdött polipeptidben a fehérjékre
jellemzô alfa helyzetû peptidkötés mellett az aszparaginsav
és a glutaminsav másik, béta vagy gamma helyzetû
karboxilcsoportja is részt vesz a peptidkötések kialakításában.
A kapott fehérjeszerû anyag katalitikus tulajdonságokkal
is bír. A fehérjék keletkezésének e
termikus útját az alkalmazott, viszonylag magas hômérséklet
miatt sok kritika érte, azonban ha meggondoljuk, hogy az ôsi
vulkánok oldalán, illetve a meteorbecsapódások
közelében uralkodó hômérséklet könnyen
elérhette a szintézishez szükséges értéket,
és ha azt is figyelembe vesszük, hogy bizonyos adalékok
alkalmazásával az alkalmazott hômérséklet
100 fok alá szorítható, a Fox-féle proteinoid
szintézis az ôsi, földi körülmények
között is elképzelhetôvé válik [5,9].
A fenti folyamatokkal párhuzamosan, vagy – kevésbé
valószínûleg – azok elôtt természetesen
felvetôdhet a ribonukleinsavak által katalizált fehérjeszintézis
lehetôsége is [13].
A fehérjék lehetséges abiotikus kialakulásának felvázolása után az enzimek evolúciójáról teszek néhány megjegyzést. Elôre szeretném azonban bocsátani, hogy az archaikus enzimek vázolandó sajátságai zömükben függetlenek attól, hogy a hordozó fehérjék a fenti abiotikus úton, avagy az elsô primitív élôlények által szintetizáltan jöttek-e létre. A fehérjetermészetû anyagok kialakulása során megjelenô elsô katalizátorok, enzimek meglehetôsen rossz hatásfokkal és igen kis specificitással mûködhettek [17]. Ez segítette az ôsi élô szervezeteket, hiszen így igen kevés számú katalizátor elegendô volt a meggyorsítandó folyamatok teljes körének viszonylagos felpörgetésére. E kezdeti enzimek hatókörét tovább növelte, hogy a folyamatokban részt vevô koenzimeknek (pl. NAD, ATP stb.) is széles tárházát köthették meg, viszonylag kis preferenciát mutatva egyikük vagy másikuk iránt [18].
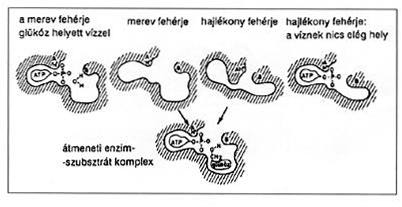 |
| 2. ábra. A glükózból ATP segítségével
glükóz-6-foszfátot képzô glükokináz enzim szelektivitása a glükóz és a tízezerszeres feleslegben lévô vízmolekula között merev és hajlékony flexibilis enzimszerkezet esetén. A primitívebb, merev szerkezetû enzim kevéssé tud különbséget tenni a glükóz és a víz között, aminek eredményeként az enzim glükokináz helyett inkább ATP-t hasító ATP-ázként mûködik |
Ezen ôsi protoenzimek viszonylag csekély katalitikus aktivitása a fehérjeszerkezet kis flexibilitásával járhatott együtt. A stabil, merev szerkezet önmagában is okozója lehetett a különbözô lehetséges szubsztrátok közötti kis szelektivitásnak (2. ábra). A fehérjeszerkezet merevségének egyik fontos oka az, hogy az adott fehérje energiája betekert (stabil) állapotában igen alacsony. Azoknak a fehérjeszerkezeteknek a száma, amelyek merevek és adott formájukba gyorsan betekeredni és e formában stabilan megmaradni képesek, igen kevés. Az ilyen fehérjéknek viszonylag kisméretûeknek kell lenniük. Ezen ôsi fehérjeformák még ma is a létezô fehérjék stabil szerkezetének jelentôs részét alkotják [19,20]. A fehérjék formavilágának magja tehát szegényes, és elképzelhetô, hogy az elsô, ôsi fehérjék alakját ôrzi ma is. Ez az igen meglepô állítás azért olyan újkeletû, mert néhány évvel ezelôttig az ismert térszerkezettel bíró fehérjék száma nem volt elég nagy ahhoz, hogy ilyen általánosító megállapításokat elég nagy biztonsággal meg lehessen kockáztatni.
Ha az elsô fehérjék kicsik, merevek, "lusták" és "buták" voltak, hogyan fejlôdött ki a mai nagy, fürge, hajlékony, "okosan" válogató fehérjék gazdag tárháza? Természetesen buta és lusta ôseikbôl. Mint már a fentiekbôl is kiderült, a mai fehérjék szerkezetének a vázát is az ôsi, merev elemek képezik. Ezekhez azonban hozzáépültek olyan hajlékony, rugalmas fehérjerészek, amelyek éppen a stabil elemek betekeredésébôl származó energiát használják ki arra, hogy saját, nem olyan stabil szerkezetük az adott fehérjében mindig közel azonos módon újraképzôdjön. Vannak-e a késôbbi fehérjerészek közül olyanok, amelyek az egész fehérje betekeredése során "mindig a rövidebbet húzzák" (övék marad a Fekete Péter) és igen labilis állapotban kell maradniuk ahhoz, hogy a fehérje többi része stabil legyen? Az ilyen részeket "aktív helyeknek" hívjuk. Az aktív helyek csak más molekula kötésével képesek stabilizálódni. Ez a más molekula lehet kicsi, ilyenkor az aktív hely egy enzim szubsztrátkötô helyévé válik, és lehet nagy is, ilyenkor az aktív hely a fehérje más fehérjéhez vagy nukleinsavhoz való kötôdését segíti elô. A fehérjék aktív helyei tehát viszonylag késôbb alakultak ki, és az enzimhatás, a makromolekulák szelektív felismerôképessége a mai élôvilág mûködése szempontjából alapvetô fontosságúak.
Mi tette lehetôvé az aktív helyek kialakulását? Egy ilyen, instabil részekkel rendelkezô fehérje csak lassan tud betekeredni [20]. Szerkezetének kellô sebességû kialakulásához olyan segítségre van szüksége, ami e betekeredést felgyorsítja. A tekeredést segítô fehérjetermészetû anyagokat molekuláris chaperonoknak hívjuk [21]. A molekuláris chaperonok megjelenése tehát a földi evolúció viszonylag korai és igen fontos eseménye kellett, hogy legyen [22]. Egy kérdéssel továbbmehetünk: mi tette lehetôvé a molekuláris chaperonok kialakulását? A chaperonok, azaz a fehérjekötô fehérjék kialakulása nem olyan nagy csoda, mint ahogy azt esetleg elsôre gondolnánk. Monomerek spontán polimerizációja esetén szinte törvényszerû, hogy a keletkezô polimer kösse azokat az anyagokat, amelyek a polimerizáció során a közelében voltak [23]. Fehérjéknek fehérjék jelenlétében történô polimerizációja esetén a keletkezô új fehérje a keletkezése során jelenlévô régebbi fehérjéket kisebb vagy nagyobb affinitással kötni képes. A molekuláris chaperonok keletkezésének magva tehát adott.
Az enzimfejlôdés egy késôbbi szakaszában, a fehérjék méretének fokozatos növekedése során kerülhetett sor a több fehérjeláncból, alegységbôl álló enzimek kialakulására. Az összetapadt fehérjeláncokból álló enzimkomplexeket nehezebben veszíti el a minden bizonnyal a mainál sokkal nagyobb áteresztôképességû határoló membránnal rendelkezô, ôsi sejt. A nagyobb enzimkomplexek lebontással szembeni ellenállóképessége is nagyobb [17].
A ribonukleinsavak ôsi világa
Bizonyos feltételezések szerint a fehérjék,
enzimek fenti világa elôtt a földi molekuláris
evolúció a ribonukleinsavak (RNS-ek) evolúciójaként
terjeszkedett. Az RNS azért annyira vonzó mint ôsmolekula,
mert egyesíteni tudja az enzimekre jellemzô katalitikus aktivitást
a megkettôzôdés (önmásolás) precizitásával,
amely mindenfajta molekuláris szintû információ
pontos megôrzésének alapvetô feltétele.
E kezdetleges világra azonban korántsem biztos, hogy az információ
továbbadásának, az öröklôdésnek
olyan molekulárisan konzervált formái voltak a jellemzôek,
mint a dezoxiribonukleinsav (DNS) másolásának pontos,
mai mechanizmusa. Az információs egységek molekulák
feletti szinteken, katalitikus körfolyamatok újra- és
újraszervezôdésében, illetve stabil molekulatársulásokban
ölthettek testet (nem egy esetben valamely szilárd felszínhez
mint további rendezôfelülethez kötötten [10,11]).
Másrészt pedig még nem is volt szükség
a létezô szervezôdések pontos öröklôdésére,
mert a meglévô formák csekély bonyolultságuk
és igen kezdetleges hatékonyságú mûködésük
miatt "kis evolúciós értéket" képviseltek.
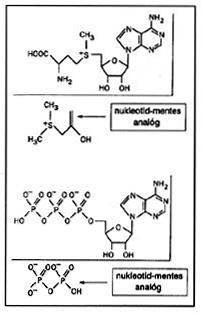 |
| 3. ábra. Különbözô koenzimek (S-adenozil-metionin és adenozin-trifoszfát, ATPT) és nukleotidmentes analógjaik |
A ribonukleinsavak fehérjék elôtti létét az is valószínûsíti, hogy a számos enzim katalitikus aktivitásában részt vevô koenzim (NAD, ATP stb.; ld. 3. ábra) az ôsi RNS-enzimek leszármazottjának is tekinthetô [5]. A legegyszerûbb RNS-enzim (ún. ribozim) pusztán hat nukleotidból, három uracilból és három adeninbôl áll (UUUAAA) [24]. Az RNS-enzimek képesek az aminosavak kötésére, és mind a fehérje-, mind pedig az RNS-szintézis primitív katalízisére [1,13,24,25]. Ugyanakkor igen sok érv szól az ellen, hogy az RNS a földtörténet közel 4 milliárd évvel ezelôtti korszakában egyeduralkodó szerves molekula lett volna: a ribóz az ôslevesnek csak kis mennyiségben jelen lévô komponense volt, ami csökkenti a tisztán ribózalapú nukleinsavak kialakulásának esélyeit. További ellenérv, hogy a keletkezô RNS nem elég stabil és polimerizációja során különbözô térállású formái összegabalyodhatnak. Így számos olyan elképzelés született, amely az RNS szerkezetében szereplô ribózt valamely más, egyszerûbb molekulával helyettesítette. Sajnos azonban ezeknek az ôsi RNS-eknek a katalitikus aktivitására mind a mai napig nincs elegendô adat [1,13].
A ribonukleinsavak fenti katalitikus aktivitását azonban nem szabad túlértékelnünk. A húszfajta különbözô aminosavból felépülô fehérjéknek becslések szerint több mint 500-szor nagyobb esélyük van egy aktív, katalitikus hely kialakítására, mint az azonos méretû, négyféle nukleotidból felépülô RNS-enzimeknek [5]. Az RNS tekeredése sokkal kevésbé kooperatív, mint a fehérjéké, így sokkal kisebb a valószínûsége annak, hogy az RNS-molekula valamely részlete kényszerûen magas energiaállapotban maradjon [26]. Ha pedig nincs lokálisan igen magas energiaállapotú RNS-darabka, akkor nincs jól funkcionáló RNS aktív hely se, vagyis az RNS-ek elvileg sem lehetnek igen hatékony enzimek. A fejlôdés egy késôbbi fokán tehát a fehérjetermészetû enzimeknek szükségszerûen teret kellett hódítaniuk. Azok a fehérjék, amelyek bizonyos RNS-enzimeket teljesen helyettesíteni képesek [27], mind a mai napig ôrzik e térhódítás emlékét. Természetesen a fehérjék messze nem minden katalitikus funkciót vettek át, jól mûködô RNS-enzimekre a ma élô szervezetekben is számos példa hozható.
Fehérje-ribonukleinsav komplexek
A fentiekben vázolt fehérje-ribonukleinsav tyúk–tojás
problémának van egy igen szellemes megoldása is: se
a fehérjék nem elôzték meg a ribonukleinsavakat,
se a ribonukleinsavak a fehérjéket. A két molekulafajta
egymás mellett, egymással kölcsönhatásban
fejlôdött ki [28]. Ezen elmélet szerint a kezdeti szakaszban
mind a fehérjék, mind pedig a ribonukleinsavak (illetve nem
ribózalapú ôseik) spontán polimerizációja
során a másik molekulafajta gyakorta jelen volt. Így
a kezdeti fehérjék RNS-t, a kezdeti RNS-ek pedig fehérjéket
kötöttek [23]. A két molekulatípus stabilizálta
egymást és egymás polimerizációjában
és betekeredésében is kölcsönösen gyorsító
szerepet tölthetett be [29].
A fejlôdés késôbbi szakaszában az RNS-ek bázispárosodásában egyre kisebb lett a "lötyögés", ezzel párhuzamosan a lehetséges nukleotidok száma négyre csökkent [5]. A csökkenô nukleotidszámmal az RNS-ek katalitikus aktivitása jelentôsen megcsappant. A kezdetben csak néhány (4–6)-féle aminosavból álló kicsi fehérjék pedig egyre újabb aminosavakat kezdtek magukba építeni és ezzel méretük is egyre nôtt. Mindezzel párhuzamosan katalitikus hatékonyságuk nagyságrendekkel javult. A kezdetben együtthaladó ôs-RNS és ôsfehérje útja az evolúció során kettéágazott: az RNS a pontos másolásra, a fehérje a katalízisre szakosodott molekulává vált. A fenti, szimpatikus kép kísérletes bizonyításához a pl. alanin-aszparaginsav típusú ôsfehérjék saját magukat lemásoló (replikáló) hatásának bizonyítása még hiányzik.
A régi fehérjeszerkezet lehetséges ôrzôi
Találhatunk-e olyan fehérjéket, amelyek az ôsi
(merev, lusta és buta) fehérjék bizonyos tulajdonságait
mind a mai napig megôrizték? A fentiek szerint ezeknek a tanúfehérjéknek
igen rossz katalitikus aktivitással kell bírniuk, igen sokféle
szubsztrátot kell kötniük, elég merev, stabil szerkezettel
kell rendelkezniük és az sem árt, ha RNS kötésére
is képesek. Szétnézve a fehérjék között
a glikolízis bizonyos ôsi enzimjei mellett a molekuláris
chaperonoknak a fehérjeszerkezet kialakulását segítô
családja az, amely a fenti kritériumoknak megfelel [21].
Elképzelhetô tehát, hogy ezek az evolúció
korai szakaszában megjelenô [22], igen konzervált szerkezettel
bíró fehérjék e korai szakaszra jellemzô
fehérjetulajdonságok némelyikét meglehetôs
változatlansággal mind a mai napig hordozzák.
Lépések az elsô sejt kialakulása felé: DNS, genetikai kód, és sejtmembrán
RNS és DNS
A nukleinsavak (RNS és DNS) egymáshoz viszonyított
evolúcióbeli sorrendjét alapvetôen stabilitási
problémák határozták meg. A monomerként
létezô ribóz sokkal stabilabb, mint a dezoxiribóz,
így a DNS-nek az RNS elôtti keletkezése felettébb
valószínûtlen. Ugyanakkor az RNS láncába
beépült ribóz éppen a DNS-be épült
dezoxiribózból hiányzó hidroxilcsoportjával
képes az RNS szál hidrolízisét (vízmolekulák
által történô hasítását)
katalizálni. Így érthetô, ha egyszer mód
nyílt a DNS kialakulására az evolúció
során, miért ôrzôdött meg e stabilabb molekulafajta.
Bizonyos vírusok mind a mai napig RNS formájában ôrzik
információs anyagukat, de ezekben a szervezôdésekben
az RNS szoros fehérjeburokba préselve védve van a
láncszakító vízmolekulák támadása
elôl [5,10,30]. A DNS-ben az RNS uraciljét helyettesítô
timin is nagyobb stabilitást kölcsönöz a DNS-molekulának
[5].
Az RNS DNS elôtti szerepére utal, hogy a ma élô szervezetekben a dezoxiribonukleotidok szintézise mindig a megfelelô ribonukleotidok redukciójával valósul meg, s a DNS megkettôzôdése során RNS-primer jelenléte szükséges. A DNS kettôs hélixet alkotó térszerkezete a dezoxiribóz hiányzó hidroxilcsoportja miatt jóval kevesebb számú térbeli szerkezetet megvalósító formában létezhet, mint az RNS hajtûket, csomókat és számos más alakzatot felvevô szerkezete. Emiatt a katalitikus tulajdonságokkal nem rendelkezô DNS az információ tárolásának sokkal kevésbé sokrétû, de megbízhatóbb eszköze, mint az RNS [5,10].
A genetikai kód kialakulása
A sejten belüli információáramlás
mai, roppantul bonyolult módját sokáig igen nehéz
volt elképzelni. A valószínûbb elképzelések
megleléséhez itt is kompromisszumokat kellett kötni.
Be kellett látni, hogy az ôsi "fehérjeszintézis"
messze nem az volt, mint a mai. Melyek lehettek a különbségek?
Jóval kevesebb, akár a jelenlegi 20 helyett csak négy-,
hatféle aminosavból állhattak a kezdeti fehérjék.
Az egyes aminosavakat kódoló nukleotidsorrend nem volt olyan
szigorúan meghatározva, mint ma. Az aminosavak enzimek segítsége
nélkül, közvetlenül kötôdhettek az ôket
szállító transzfer-RNS-ekhez. Végezetül:
a bonyolult riboszómák helyett RNS-enzimek végezhették
el az ôsi, kis hatékonyságú fehérjeszintézist.
A kevésbé bonyolult, ôsi genetikai kódokat a
meglévô kód számos tulajdonsága még
ma is ôrzi (az elsô nukleotid a kódolt aminosav nagyságára
jellemzô: purin/kicsi, pirimidin/nagy; a második nukleotid
utal arra, hogy a kódolt aminosav hol helyezkedik el: purin/aminosav
a fehérje külsején, pirimidin/aminosav a fehérje
belsejében; a harmadik nukleotid pedig sokszor nem teljesen meghatározott,
"lötyög"; 4. ábra [5].)
| 4. ábra. A sejtmagbeli genetikus kód |
|||||||
| Kód | Aminosav | Kód | Aminosav | Kód | Aminosav | Kód | Aminosav |
| UUU | Phe | UCU | Ser | UAU | Tyr | UGU | Cys |
| UUC | Phe | UCC | Ser | UAC | Tyr | UGC | Cys |
| UUA | Leu | UCA | Ser | UAA | Ter | UGA | Ter |
| UUG | Leu | UCG | Ser | UAG | Ter | UGG | Trp |
| CUU | Leu | CCU | Pro | CAU | His | CGU | Arg |
| CUC | Leu | CCC | Pro | CAC | His | CGC | Arg |
| CUA | Leu | CCA | Pro | CAA | Gln | CGA | Arg |
| CUG | Leu | CCG | Pro | CAG | Gln | CGG | Arg |
| AUU | Ile | ACU | Thr | AAU | Asn | AGU | Ser |
| AUC | Ile | ACC | Thr | AAC | Asn | AGC | Ser |
| AUA | Ile | ACA | Thr | AAA | Lys | AGA | Arg |
| AUG | Met | ACG | Thr | AAG | Lys | AGG | Arg |
| GUU | Val | GCU | Ala | GAU | Asp | GGU | Gly |
| GUC | Val | GCC | Ala | GAC | Asp | GGC | Gly |
| GUA | Val | GCA | Ala | GAA | Glu | GGA | Gly |
| GUG | Val | GCG | Ala | GAG | Glu | GGG | Gly |
Sejtmembrán
Ahhoz, hogy egy ôsi sejt sejt legyen, el kellett különítenie
magát a környezetétôl. Az elkülönüléshez
hosszú ideig elegendô lehetett az, hogy a sejtszerû
szervezôdésre jellemzô kémiai folyamatok valamilyen
ásvány felszínéhez tapadva zajlottak [10,11].
Egy idô után azonban a kétdimenziós lehetôségek
szûknek bizonyultak, és a (minden bizonnyal pozitív
töltésû) felszínhez kötött ôsi
fehérjék, nukleinsavak kezdtek leszakadozni. A térbeli
elzártság megteremtésére olyan lipidszerû
anyagok adtak lehetôséget, amelyek önmaguktól
a belsô teret határoló membránná képesek
összeállni. Az elsô membránok még bizonyára
az ásványi felszínhez tapadt, lyukas hártyák
lehettek. Késôbb alakultak ki azok a mechanizmusok, amelyek
a membránokon bizonyos töltéssel rendelkezô anyagokat
(pl. egyszerû ionokat) szelektíven átjuttattak. Ekkor
szakadhattak el a membránok véglegesen az ásványi
felszínektôl. A membránnal körülvett sejtszerû
struktúrák osztódása pedig abban az esetben,
ha a szervezôdésnek egy meghatározott felszín/térfogat
arányt tartania kell, törvényszerû folyamat [5].
Eszmefuttatásunkban eljutottunk tehát az egyszerû, szervetlen molekuláktól az információhordozó, anyagcserével rendelkezô, osztódó sejtig. A lehetséges folyamatokat végigkövetô okfejtés botladozó és helyenként bizony lyukas. Még ilyen hiányos változatának végiggondolása is nagyon felemelô leckéje lehet azonban annak, hogyan képes az emberi gondolkodás és kísérletezôkedv elemi röghözkötöttségein, a cikk elsô részében említett idô- és térbeli korlátain felülemelkedni.
Ajánlott irodalom
[1] Schidlowski, M. (1988) A 3,800-million-year isotopic record of life from carbon in sedimentary rocks. Nature 333, 313-318; Lazcano, A. and Miller, S.L. (1996) The origin and early evolution of life: prebiotic chemistry, the pre-RNA world, and time. Cell 85, 793-798
[2] Crick, F.H.C.: Az élet mikéntje. Gondolat, Budapest, 1987; Gánti, T.: Kontra Crick, avagy Az élet mivolta. Gondolat, Budapest, 1989
[3] McKay, D.S., Gibson, E.K.Jr., Thomas-Keptra, K.L., Vali, H., Romanek, C.S., Clement, S.J., Chillier, X.D.F., Maechling, C.R. and Zare, R.N. (1996) Search for past life on Mars: possible relic biogenic activity in martian meteorite ALH84001. Science 273, 924-930
[4] Gánti Tibor: Az élet eredete -- újabb kutatások és eredmények. A biológia aktuális problémái sorozat (szerk.: Csaba György) 1. kötet 45-90 oldal, Medicina, Budapest, 1974
[5] Maynard Smith, J. and Szathmáry, E.: The major transitions in evolution. W.H. Freeman, 1995; Szathmáry E. and Maynard Smith, J. (1995) The major evolutionary transitions. Nature 374, 227-232; Szathmáry, E. és Maynard Smith, J. (1995) A genetikai rendszerek eredete. Természet Világa 126/1. különszám 20-26
[6] Eigen, M., Schuster, P., Gardiner, W and Winkler-Oswatitsch, R. (1981) The origin of genetic information. Scientific American 244, 78-94
[7] Miller, S.L. (1953) A production of amino acids under possible primitive Earth conditions. Science 117, 528-529
[8] Chyba, C. and Sagan, C. (1992) Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: an inventory for the origins of life. Nature 355, 125-132
[9] Kajtár Márton (1963) Vitális jelentôségû szénvegyületek keletkezésének lehetôsége a Földön az élet megjelenése elôtt. MTA Kémiai Oszt. Közl. 20: 1-32
[10] Wächtershäuser, G. (1988) Before enzymes and templates: theory of surface metabolism. Microbiol. Rev. 52, 452-484
[11] Wächtershäuser, G. (1992) Groundworks for an evolutionary biochemistry. The iron-sulphur world. Prog. Biophys. Molec. Biol. 58, 85-201
[12] Fox, S.W. (1965) A theory of macromolecular and cellular origins. Nature 205, 328-340
[13] Joyce, G.F. (1989) RNA evolution and origins of life. Nature 338, 217-224
[14] Cooper, A.A. and Stevens, T.H. (1995) Protein splicing: self-splicing of genetically mobile elements at the protein level. Trends in Biochem. Sci. 20, 351-356
[15] Lee, D.H., Granja, J.R., Martinez, J.A., Severin, K. and Ghadiri, M.R. (1996) A self-replicating peptide. Nature 382, 525-528
[16] Matthews, C.N. and Moser, R.E. (1967) Peptide synthesis from hydrogen cyanide and water. Nature 215, 1230-1234
[17] Koshland, D.E. Jr. (1976) The evolution of function in enzymes. Fed. Proc. 35, 2104-2111
[18] Kacser, H. and Beeby, R. (1984) Evolution of catalytic proteins or On the origin of enzyme species by means of natural selection. J. Mol. Evol. 20, 38-51
[19] Li, H., Helling, R., Tang, C and Wingreen, N. (1996) Emergence of preferred structures in a simple model of protein folding. Science 273, 666-669
[20] Finkelstein, A. (1994) Implications of the random characteristics of protein sequences for their three-dimensional structure. Curr. Op. Struct. Biol. 4, 422-428; Privalov, P.L. (1996) Intermediate states in protein folding. J. Mol. Biol. 258, 707-725.
[21] Csermely, P. (1995) Napjaink stressze -- sejtjeinkben (avagy: mit tesz a sejt ha bántják?) Természet Világa 126, 318-320
[22] Todd, M.J., Lorimer, G.H. and Thirumalai, D. (1996) Chaperonin-facilitated protein folding: yield by an iterative annealing mechanism. Proc. Natl. Acad. Sci. USA 93, 4030-4035
[23] Pande, V.S., Grosberg, A.Y. and Tanaka, T. (1994) Thermodynamic procedure to synthesize heteropolymers that can renature to recognize a given target molecule. Proc. Natl. Acad. Sci. USA 91, 12976-12979
[24] Pyle, A.M. (1993) Ribozymes: a distinct class of metalloenzymes. Science 261, 709-714
[25] Czech, T.R. (1986) A model for the RNAócatalyzed replication of RNA. Proc. Natl. Acad. Sci. USA 83, 4360-4364; Ekland, E.H. and Bartel, D.P. (1996) RNA-catalysed RNA polymerization using nucleoside triphosphates. Nature 382, 373-376
[26] Draper, D.E. (1996) Strategies for RNA folding. Trends in Biochem. Sci. 21, 145-149
[27] Mohr, G., Caprara, M.G., Guo, Q. and Lambowitz, A.m. (1994) A tyrosyl-tRNA synthetase can function similarly to an RNA structure in the Tetrahymena ribozyme. Nature 370, 147-150
[28] Lahav, N. (1991) Prebiotic co-evolution of self-replication and translation or RNA world? J. Theor. Biol. 151, 531-539
[29] Woodson, S.A. (1996) How RNA and protein get together. Curr. Biol. 6, 23-25
[30] Westheimer, F.H. (1987) Why nature chose phosphates. Science 235, 1173-1178
| Természet Világa, | 128. évf. 1. sz. 1997. január, 10–14. o. |
| https://www.kfki.hu/chemonet/TermVil/ | |
| https://www.ch.bme.hu/chemonet/TermVil/ |