Molnár Gergely, Blaha Béla, Horváth
Gábor
Látás az ibolyán túl
Az ultraviola fény érzékelésének
elterjedése és szerepe az állatvilágban
Viszonylag régóta közismert már, hogy számos
rovarfaj érzékelni tudja az ultraviola (UV) fényt,
ugyanakkor az, hogy erre a gerinces állatok is képesek, kuriózumnak
számított, s szerepét legtöbbször homály
fedte. Az elmúlt években azonban kiderült, hogy az ultraibolya
fotoreceptorok igen elterjedtek az állatvilágban, és
egyre inkább az számít kivétenek, ha valamelyik
magasabb rendû állatcsoportban még nem leltek UV-érzékeny
fajra. Lassan tisztázódni látszik az ibolyántúli
fény érzékelésének a tájékozódásban,
a táplálékszerzésben, a fajon belüli kommunikációban
és a bioritmus szabályozásában betöltött
fontos szerepe. Az állatok UV-érzékelésének
vizsgálata a látáskutatás egyik új és
rohamosan fejlôdô ága, amely még évekre
munkát ad a kutatóknak, hiszen annyi még a megoldatlan
biológiai és érzék-biofizikai kérdés,
s oly sok az UV-érzékelés szempontjából
eddig nem vizsgált fajok száma. Feltehetôen hatással
van és lesz az "ózonlyuk"-jelenség is az
élôvilágra, ami segítheti az UV-mintázattal
és UV-látással bíró fajok elterjedését.
Cikkünkben az emberi szem számára láthatatlan
UV-fény érzékelésének az állatvilágbeli
elôfordulását és szerepét tekintjük
át. Néhány kép erejéig azt is bemutatjuk,
miként néznek ki egyes virágok és lepkék
az ibolyántúli fényben. (Az írás OTKA-pályázatok
támogatásával készült.)
A legtöbb gerinctelen és gerinces faj érzékelni
képes az ultraibolya fényt is (1.a)-d) ábra),
amit mi emberek nem (1.e) ábra). Ahhoz azonban, hogy megérthessük
ezen állatok látás által irányított
magatartását, meg kell vizsgálnunk azokat a környezeti
vizuális jeleket, amelyekre az állatok válaszolnak.
 |
 |
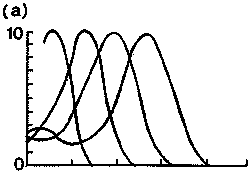
1. ábra. Különbözõ fajok
látópigmentjeinek elnyelési spektruma. Függõleges
tengely:
relatív abszorpció; vízszintes tengely: nanométerben
mért hullámhossz.
a) lepke négy vizuális pigmenttel (360, 437, 500, 568 nm);
b) aranyhal négy
pigmenttel (360, 432, 542, 620 nm) (Kissé módosított
ábra [5] alapján) |
Az emberi szem a 400 nm-tõl 800 nm-ig terjedõ hullámhosszúságú
fényt (ez a látható vagy optikai tartomány)
érzékeli (1.e) ábra). A 200 nm-tõl 400
nm-ig húzódó spektrális zónát
nevezik ultraviolának (UV), amely további három részre
osztható: UV-A, B, C. A napfény UV-C (200-280 nm) összetevõjét
teljesen elnyeli a földi légkör, az UV-B-sugarakat (280-330
nm) pedig részben kiszûri a légköri ózonréteg.
Az UV-B sugárzás intenzitása azonban az ózonpajzs
elvékonyodása (“kilyukadása”) miatti növekszik,
többek között ez okozza az egyre gyakoribb bõrrákot
is.
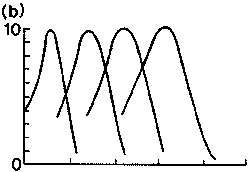
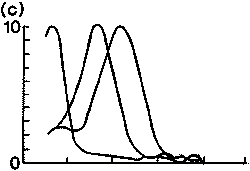 |
 |
1. ábra. Különbözõ fajok
látópigmentjeinek elnyelési spektruma. Függõleges
tengely:
relatív abszorpció; vízszintes tengely: nanométerben
mért hullámhossz.
(c) közönséges gekkó három pigmenttel (364,
467, 521 nm); (d) pekingi vörösbegy
négy pigmenttel (355, 454, 526, 600 nm) |
Az UV-A-sugárzás (320-400 nm) lejut a Föld felszínéig,
s ez az a tartomány, amit egyes UV-érzékeny szemû
állatok kiaknáznak (1.a)-d) ábra).
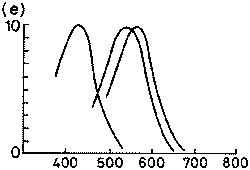 |
1. ábra. Különbözõ fajok látópigmentjeinek
elnyelési spektruma. Függõleges tengely:
relatív abszorpció; vízszintes tengely:
nanométerben mért hullámhossz.
(e) ember három pigmenttel (420, 530, 565) |
Az emberi szem szaruhártyája és lencséje
erõteljesen elnyeli a természetes UV-fényt, amely
így nem jut el a recehártyáig és nem okozhatja
annak károsodását. Mivel azonban a kék fényre
érzékeny fotoreceptorok elnyelési spektrumának
lecsengõ vége az UV-A tartományba is belenyúlik
(1.e) ábra), ha a szemlencsét pl. szürkehályog
megszüntetésének céljából mûtéttel
mûanyag lencsével cserélik ki (ez már átengedi
az UV-fényt), akkor a páciens már képes az
ultraibolya fény részleges érzékelésére.
Az UV-fény persze az ilyen ember számára nem “ibolyántúli
színekben”, hanem kékben/ibolyában jelenik meg. Mivel
sok szerves molekula (pl. DNS) elnyeli az UV-fényt, a még
viszonylag nagyobb, 380 nm körüli hullámhosszúságú
UV-fény is a retina sérülését eredményezi,
és rákot okozhat. Ezért számunkra az a jó,
ha szemünk dioptrikus készüléke kiszûri a
káros UV-fényt. Ezen elnyelés hatására
a lencse anyaga különbözõ, számunkra is látható
színekben fluoreszkál.
Az állatok zöme viszont közvetlenül érzékeli
az ultraibolya fényt. Némelyikük erre a célra
speciális UV-fotoreceptorokat fejlesztett ki (1.a)-d)
ábra). Mások viszont az UV-sugarakat áteresztõ
dioptrikus készülékkel bírnak. Ez utóbbiak
az UV-fényt a kék fényre érzékeny fotoreceptoraikkal
detektálják, amelyek elnyelési spektruma belenyúlik
az UV-tartományba. E két eltérõ vizuális
stratégia elõnyei és hátrányai még
nem kellõen tisztázottak.
Az, hogy egy állatnak több, különféle színre
érzékeny fotoreceptora van, még nem jelenti azt, hogy
ezeket az állat színlátásra használja.
A több, eltérõ látópigment alkalmazását
az is indokolhatja, hogy az állat a spektrum szélesebb tartományában
legyen képes a fényt érzékelni és így
jól lásson a nap bármely szakában. Nappal pl.
a közvetlen napfény (melynek intenzitásmaximuma a spektrum
zöldessárga tartományába esik) dominál.
Ekkor a közepes hullámhosszakra érzékeny csapok
használata az elõnyös. Naplemente, ill. napfelkelte
idején ellenben a vöröses-narancssárgás
napsugarak által megvilágított, hosszabb hullámhosszakra
érzékeny fotoreceptorok az optimálisak. Ha
ekkor egy bizonyos viselkedési mintát - pl. rajzást
(kérészek) vagy vándorlást (madarak) - a kék
és ultraibolya összetevõben gazdag szórt égboltfény
vezérel, akkor a rövid hullámhosszakra érzékeny
fotoreceptorok alkalmazása a kedvezõ. Színlátás,
azaz a fény eltérõ hullámhosszainak a megkülönböztetése
csak akkor történhet, ha az eltérõ csapokban
kiváltott elektromos jeleket a neurális hálózat
összehasonlítja, s az adott hullámhosszat a fotoreceptorok
elektromos aktivitásainak a viszonyai kódolják. Tehát
a színlátáshoz szükséges, de nem elégséges
feltétel a különbözõ fotoreceptorok megléte.
Az UV-látás fajok szerinti eloszlása
Sok gerinctelennek van UV-receptora; ebbõl a szempontból
a legrészletesebben a rovarokat és a rákokat vizsgálták.
A rovarok képviselõinek retinájában
rendre kettõ (di), három (tri), ill. négy (tetra)
különbözõ látópigment fordul
elõ. Habár a pigmentek száma változó,
a legtöbb rovarnak van UV-receptora is (1.a) ábra),
amelynek átlagban 336 nm-en van a legnagyobb érzékenysége.
A fentiek alapján az UV-pigmenteknek a retinában való
jelenléte persze nem jelenti feltétlenül azt, hogy azok
a színlátást szolgálják. Néhány
rovarnál azonban viselkedéstani kísérletekkel
kimutatták, hogy képesek a színeket megkülönböztetni
az UV-tartományban. A legintenzívebben tanulmányozott
rovar a mézelõ méh, amelynek három eltérõ
fotoreceptor-csoportja van: 340 nm (UV), 430 nm (kék) és
535 nm (zöld). E spektrális tartományokban a méh
jól meg tudja különböztetni a hullámhosszakat,
jelezve, hogy az UV-receptorok a színlátó rendszerbe
teljesen beilleszkedtek.
A rákok többsége a rovarokénál
kevésbé fejlett színlátással rendelkezik;
gyakran csak két fotopigmenttel, de ezek egyike általában
UV-érzékeny. A színlátásban a rákok
között a sáskarákok a csúcstartók,
amelyeknek 10-11 látópigmentjük van. Ha figyelembe vesszük
a retinabeli különbözõ színszûrõ
cseppeket is, akkor 16 különbözõ fotoreceptor-osztályt
kapunk; közülük hat 300-400 nm között nyeli el
a fényt. Ez arra utal, hogy az UV-tartományban rendkívül
jó a sáskarákok színlátása.
Nagyon sok halfajnál kimutatták már az UV-receptorok
jelenlétét (1.b) ábra). Ezekben a 360-365 nm
közé esõ elnyelési maximummal bíró
UV-pigmentek a retina elkülönült rövid csapjaiban találhatóak,
amelyek pl. a pisztrángoknál egy négyzetes retinális
hálózat csomópontjaiban helyezkednek el. E halakban
az UV-csapok nemcsak egyszerû UV-detektorok, hanem a színlátásban
is részt vesznek. Az aranyhalaknak pl. négy csappigmentjük
van az UV-val (360 nm) együtt, és a viselkedéstani kísérletekben
a spektrális érzékenységi függvényüknek
az alakja tisztán mutatja a színlátásra utaló
csúcsokat és völgyeket.
Az UV-receptorok szempontjából eddig csak kevés
kétéltû és hüllõ retináját
tanulmányozták. A kétéltûek közül
két szalamandra fajban mutatták ki eddig az UV-érzékeny
pigment jelenlétét. A közönséges szalamandra
pigmentje elnyelési spektrumának a maximuma a kék
tartományban (420 nm) van, és csak egy része nyúlik
át az UV-sávba. A hüllõknél eddig nyolc
fajban találtak UV-receptorokat: egy teknõsben, két
gekkófajban (1.c) ábra) és öt gyíkban.
Ezek az UV-csapok az összes csapnak csak néhány százalékát
teszik ki. A viselkedéstani kísérletek szerint az
említett teknõs az UV-receptorait színlátásra
használja.
A madaraknál eddig harmincnál több faj szeme
bizonyult UV-érzékenynek. Általában rendkívül
jó színlátásúak, s legtöbbjük
négy-öt csappigmenttel is rendelkezik. Ezek közül
az egyik vagy az UV-tartományban nyeli el a fényt a legerõsebben
(pl. pekingi vörösbegy, 1.d) ábra), vagy a látható
tartomány rövid hullámhosszára a legérzékenyebb,
és az abszorpciós függvény kiterjed az UV-tartományra
is (pl. a tõkés récében). Ezen UV-receptorok
teljesen a madár színlátó rendszerébe
integrálódtak, amit számos madárfaj spektrális
érzékenységi függvénye bizonyít
és sok faj azon képessége, hogy az UV-tartományban
is meg tudja különböztetni az eltérõ hullámhosszakat.
Sok madár retinájában találtak már ún.
dupla csapokat, amelyek morfológiailag eltérõ fõ
és járulékos tagból állnak. Az eddig
vizsgált fajokban csak egyszerû csapokban találtak
UV-pigmenteket, de ezek a csapok ritkábbak és kisebbek is,
mint a többi egyszerû csapsejt.
A madarak látásának kutatásában igen
kedvelt faj a pekingi vörösbegy. E madár szemének
optikai elemei jól áteresztik a 400 nm alatti hullámhosszúságú
fényt, s ezt erõteljesebben csak 330 nm-tõl lefelé
terjedõ tartományban kezdik elnyelni. A retinában
található csapsejtek külsõ szegmensében
különbözõ színû olajcseppecskék
helyezkednek el, amelyek megszûrik a beérkezõ fényt,
mielõtt az a fotopigmenteket eléri. A pekingi vörösbeggyel
végzett viselkedéstani kísérletek során,
a 300-400 nm között a hullámhossz függvényében
felvett, fénnyel szemben mutatott relatív érzékenységi
görbe négy, egymástól élesen elhatárolódó
maximumot mutat a 370, 460, 530 és a 620 nm-es értékeknél.
A négy érzékenységi maximumnak megfelelõen
négy, egymástól fotopigmentjeikben és olajcseppecskéikben
eltérõ csapsejtet találtak a retinában. A nagyobb
hullámhosszakra érzékeny csapok (568, 499 nm) vörös,
ill. narancssárga olajcseppecskéket tartalmaznak, míg
a rövidebb hullámhosszakra érzékenyek (454, 355
nm) sárgát, ill. színtelent. Hogy e madarak mire használják
az UV-látásukat, még csak részben tisztázott.
Az mindenesetre már bizonyított, hogy a hímek UV-fényben
a nõstényekkel ellentétben feltûnõen
világos megjelenésûek, és a nõstények
elõszeretettel választanak szexuális partnerül
UV-ben világosabbnak tûnõ hímeket.
Az emlõsök a fõemlõsök kivételével
fõleg dikromatikus színlátásúak.
Egyik csaptípusuk spektrális elnyelési görbéjének
maximuma 530-560 nm között van, míg a másiké
420-440 nm közötti. Néhány, éjszaka aktív
rágcsálónál mindkét pigmenttípus
a rövidebb hullámhosszak felé tolódott el. A
házi egérben és a mongol versenyegérben
pl. a nagyobb hullámhosszakra érzékeny pigment abszorpciós
maximuma 500-510 nm közé esik, és a rövidebb hullámhosszak
érzékeléséért felelõs pigmenté
360-370 nm közé. Ez az eltolódás együtt
jár az UV-fényt áteresztõ szemlencsével.
A viselkedéstani kísérletek tanúsága
szerint az utóbbi két faj UV-receptorait a színlátásban
használja. Az emlõsök között eddig csak e
két fajnál sikerült UV-pigmentet kimutatni, habár
az emlõsök igen széles skáláját
tanulmányozták már e szempontból. Természetesen
más emlõsök is képesek lehetnek érzékelni
az UV-fényt, amennyiben szemük dioptrikus készüléke
átengedi azt, mivel a rövidebb hullámhosszak érzékeléséért
felelõs csapjuk látópigmentjének abszorpciós
spektruma az UV-tartományba is belenyúlik. Egy lehetséges
példa erre a patkány, amelynek a szemlencséje
350 nm-en még 50%-os áteresztõképességgel
bír.
Az UV-látás szerepe a tájékozódásban
A fény egy átlátszó közegen áthaladva
szóródik és részlegesen síkban polarizálódik
a közeg részecskéin. Ha ezek mérete sokkal kisebb
a fény hullámhosszánál (pl. porszemcsék,
levegõmolekulák, a levegõ sûrûségingadozásai),
akkor a szóródás és a polarizáció
mértéke a hullámhossz negyedik hatványának
a reciprokával arányos (elsõdleges Rayleigh-szóródás).
Így a rövidebb hullámhosszak, mint az UV vagy a kék,
sokkal jobban szóródnak és polarizálódnak
a hosszabbaknál. A szóródás miatti polarizáció
a beesési irányra merõlegesen a legnagyobb, ezért
az égbolton a részlegesen poláros égboltfény
elektromos terének vektorai (E-vektorok) koncentrikus köröket
képeznek a Nap körül. Ha egy állat képes
detektálni ezen E-vektorokat, akkor meg tudja állapítani
a Nap irányát az égbolt polarizációs
mintázatából még akkor is, ha a Nap nem látszik
a felhõk vagy a tereptárgyak miatt. Néhány
rovar ily módon detektálja a Nap irányát.
Eddig két rovarfaj navigációs képességeit
vizsgálták részletesen: a mézelõ
méhét és a sivatagi hangyáét.
UV-receptoraik zöme összetett szemük háti sávjában
koncentrálódik, amely az összes fotoreceptornak csak
néhány százalékát képviseli.
Ezek az UV-receptorok az ultraibolya polarizációra érzékenyek
és a térbeli tájékozódásban használatosak.
E rovarok természetesen nem végeznek semmiféle bonyolult
égi trigonometrikus számításokat, és
nem rendelkeznek az égbolt részletes térképével
sem, csak az égbolt egyszerûsített E-vektor eloszlását
ismerik, amely az UV-receptorok irányulásán alapul.
Minden UV-receptor maximálisan érzékeny egy adott
E-vektorra. A receptorok preferált E-vektor iránya folyamatosan
változik a szem háti, fejtetõi részének
elejétõl a végéig. A rovar úgy
használja belsõ E-vektor térképét,
hogy egy függõleges tengely mentén addig fordul - ez
általában a talajon történik -, amíg a
legjobban fedésbe nem kerül az ég aktuális polarizációs
mintázatával. Ekkor a fotoreceptorok maximális elektromos
jelet generálnak, ami arról tudatja a rovart, hogy éppen
szemben vagy háttal áll a Napnak. A rovar E-vektor térképe
állandó, az égbolton csak naplementekor, ill. napfelkeltekor
fellépõ E-vektor eloszlásnak felel meg, de a jobb
és bal szemben fellépõ hiba gyaníthatóan
ellenkezõ elõjelû, így kiejthetik egymást.
Sok más rovar is, mint pl. a hártyásszárnyúak,
tücskök és lepkék szemük fejtetõi
régióján a polarizáció-érzékeny
receptorok hasonló elrendezésével rendelkeznek, s
van néhány bizonyíték arra is, hogy ez az UV-érzékeny
rendszerük a fentiekhez hasonlóan mûködik.
Egyes halak szintén használják a polarizált
fényt a tájékozódáshoz. Az aranyhalakban
pl. a négy csaptípus közül a két, nagyobb
hullámhosszakra és az egy, UV-fényre érzékeny
fotoreceptor érzékeny a fénypolarizációra
is. Az UV-csapok preferált polarizációs iránya
merõleges a másik két csapéra. Az ezekbõl
eredõ elektromos jelek az alapjai annak a mechanizmusnak, amellyel
e halak meg tudják határozni a környezetükbõl
jövõ fény rezgéssíkját. Ezt az
információt orientációra használják,
ha más használható vizuális jelek, pl. fényesség
vagy szín nem állnak a rendelkezésükre. A pisztrángok
még a kevésbé poláros fény esetén
is meg tudják állapítani az E-vektor irányát
és ez alapján választják meg úszási
irányukat. E halak a fény polarizációs irányának
a vízben uralkodó viszonyai alapján képesek
vizuálisan is meghatározni a függõleges irányt,
aminek ismerete igen fontos a térbeli orientációban.
A Nap iránya (amikor nem látszik) persze nemcsak az égboltfény
polarizációja alapján határozható meg,
hanem az égboltfény erõsségének és
színének gradienseibõl is. Mivel a rövidebb hullámhosszak
jobban szóródnak a hosszabbaknál, ezért a rövidebb
hullámhosszúságú szórt fénynek
egyenletesebb az égbolton tapasztalható intenzitás-eloszlása.
A nagyobb hullámhosszúságú szórt fény
erõssége pedig rohamosan nõ a horizont felé.
Ez az oka a jól ismert horizont kivilágosodási jelenségnek.
Így az égboltról jövõ bármely két
különbözõ hullámhosszúságú
fény intenzitásának a viszonya a Naptól mért
szöggel változik. Ezért egy állat számára
az égbolt egy adott részén a spektrális összetétel
analizálása - a nagyobb hullámhosszúságú
fény egybevetése a viszonylag homogén UV-háttérrel
- érzékeny becslést szolgáltathat a Nap irányáról
még akkor is, ha a Nap nem látszik. Valószínûsíthetõen
egy ezen az elven mûködõ mechanizmus fontos szerepet
játszik bizonyos madarak tájékozódásában.
A galambokon túl más madarakról is kimutatták
már az UV-érzékenységet, és számos
vonuló énekesmadárról bebizonyították,
hogy vándorlása során az égbolt polarizációs
mintázata alapján is képes tájékozódni,
ha más érzéki jelek nem állnak a rendelkezésére.
E madarakban az UV-érzékenység szoros kapcsolatban
áll a polarizációérzékeléssel,
aminek anatómiai és neurális háttere egyelõre
ismeretlen.
Táplálékkeresés
UV-fény segítségével
Az UV-pigmenteket néhány rovar, hal, hüllõ,
madár és emlõs a színérzékelésben
használja, de a térbeli érzékelésükben
is szerepük lehet. Azok az állatok, amelyeknek látását
az UV-pigmentek is javítják, könnyebben tudják
érzékelni a táplálékukat és felismerni
fajtársaikat. Az UV-érzékelésnek a táplálékszerzésben
persze csak bizonyos körülmények között lehet
szerepe. Napközben pl. az erõs látható fény
teljesen elfedheti az UV-reflexiót, de szürkületkor vagy
borús idõben segíthet a terület felderítésében.
Galamboknál mutatták ki, hogy UV-fényben is
megkülönböztetik az eltérõ formákat,
s ebben mindkét rövidhullámhosszú receptoruk
szerepet játszik. A legtöbb gerinces fajban az UV-receptorok
száma viszonylag kevés (az összes fotoreceptor 5-10
%-a), ezért önmagában ez az UV-mozaik nem alkothat nagyfelbontású
rendszert.
Hiába is lenne sok UV-recptor a retinán, velük mégsem
lehetne élesen látni. Az ibolyántúli sugarak
ugyanis könnyen szóródnak a levegõ- ill. a vízrészecskéken,
emiatt a távoli tárgyak a légkörben vagy a vízben
UV-fényben csak homályosan látszanának. A rövidebb
hullámhosszúságú fény, így az
UV is jobban szóródik a szem optikai rendszerének
az anyagában (az ereken, buborékokon, sejteken). Ehhez adódik
még a szem törõközegének színi hibája,
azaz hogy a különbözõ színû fénysugarak
a szemlencsén eltérõ mértékben törnek
meg. (Például az emberi szemlencse fókusztávolsága
kék fényre kb. 0.5 mm-rel rövidebb, mint a pirosra).
Ha egy tárgy nagy hullámhosszúságú fényben
élesen látszik, azaz képe a retinára fókuszálódik,
akkor a kép rövid hullámhosszúságú
UV-fényben fókuszon kívülre esik, így
a tárgy életlennek, homályosnak tûnik, a szem
pedig távollátóvá válik. Ezért
tehát a széles spektrumtartományra érzékeny
fotoreceptorok alkalmazása rontaná a térbeli felbontást.
Így a nagyfelbontású optikai rendszert inkább
a szóródásra kevésbé érzékeny
nagyobb hullámhosszakat felfogó receptorokra érdemes
alapozni. Az UV-receptorok analóg módon mûködhetnek
az ember rövidhullámhosszú fotoreceptoraival: a színérzékelésben
és a durva felbontásban játszhatnak szerepet, de valószínûleg
nem vesznek részt a finom felbontású látásban,
mert az a sokkal nagyobb számban jelen levõ, nagyobb hullámhosszakra
érzékeny receptorok feladata.
Élelemkeresésre akkor használhatók UV-receptorok,
ha a táplálék a háttérhez képest
erõsen elnyeli, szórja vagy visszaveri az ibolyántúli
fényt. A sáskarákok pl. UV-fényt is
használnak táplálékuk és ellenségeik
felfedezésére, mert UV-receptoraik szélesebb spektrális
ablakot nyitnak az optikai környezetre. Halaknál elsõsorban
olyan fiatal egyedeknek vannak UV-receptoraik, amelyek a felszíni
vizekben keresik planktoni táplálékukat. A plankton
kifejezetten a rövidebb hullámhosszúságú
fényt szórja. Bizonyos halfajoknál kimutatták,
hogy UV-érzékelésük ténylegesen részt
vesz a planktonszervezetek felkutatásában, így nem
meglepõ, hogy amikor felnõnek és megváltoznak
táplálkozási szokásaik, elvesztik UV-látásukat.
Az egyéves sebes pisztrángok kezdetben még
négyféle fotoreceptorral bírnak (355, 441, 535 és
600 nm), de az elkövetkezõ életévük során
elvesztik UV-pigmentjeiket az azokat tartalmazó csapokkal együtt.
Még legalább négy fajban megfigyelhetõ ez a
jelenség, pl. a nemes lazacnál, a szivárványos
pisztrángnál, a vörösszárnyú
keszegnél és a sárga sügérnél.
E változás a hal egyedfejlõdése során
bekövetkezõ egyéb, mélyrehatóbb változásokhoz
kapcsolódik, inkább méret, mint korfüggõ.
Az UV-receptorok elvesztése egy pajzsmirigyhormon, a tiroxin
befolyása alatt áll. A tiroxin vérbeli koncentrációjának
változtatásával csökkenthetõ ill. növelhetõ
a receptorelvesztés mértéke. Sõt tiroxinkezeléssel
idõs, UV-receptorait már elvesztett halakban UV-pigment szintézise
indukálható. Az UV-receptorok arányának csökkenése
a retinában szorosan összefügg a szemlencse fényelnyelésének
növekedésével, ami a rövid hullámhosszaknál
jelentõs, s ezzel egyidõben kevesebb lesz a retinára
jutó UV-sugarak mennyisége. A lencse elnyelésének
ilyen változása általános a halak körében.
Az is lehetséges, hogy az UV-érzékenység eltûnik
az idõs példányoknál, s talán csak a
fiatal egyedekben van jelen. De vannak kivételek is: pl. a Chromis
punctipinnis nevû halfaj fiatal egyedeiben nincs UV-pigment,
csak az idõsekben.
A víz szórja a rövid hullámhosszúságú
fénysugarakat, így az UV-t is. E szórás kisebb
távolságokon belül olyan tárgyak körvonalazásában
játszhat szerepet, amelyeket esetleg más módon nehéz
lenne érzékelni. Sok hal csillogó teste azt a célt
szolgálja, hogy a háttérhez hasonló összetételû
és erõsségû fényt verjen vissza. Ez az
álca nem mûködik egy fényt szóró
és polarizáló háttér elõtt, mert
a hal pikkelyeirõl visszavert fény polarizációja
eltér a háttér szórt fényének
polarizációjától. Szintúgy, ha egy pikkelynek
a spektrum látható tartományában történõ
csillogása fényinterferencia útján alakul ki,
akkor az ibolyántúli tartományban a tükrözése
már nem olyan hatékony. Egy ilyen pikkelyekkel borított
hal jól látható a fényes UV-háttér
elõtt.
Számos, madarak által fogyasztott gyümölcs
és mag visszaveri az ultraibolya-fényt, míg
a növényi levelek nem. Hasonlóképpen sok,
a madarak potenciális prédájának számító
pillangó szárnyai UV-fényt is reflektálnak.
A virágok színe fontos információforrás
a táplálék után kutató beporzók
számára, bár a virágok alakjának is
nagy szerepe van ebben. A visszavert UV-fény tehát kulcsinger
a virágokat beporzó rovarok számára csakúgy,
mint egyes madaraknak (pl. kolibriknek).
 |
2. ábra. Egy fiatal napraforgó virágzata
ilyennek látszik UV-fényben. A felvétel
során a növényt UV-fényt is kibocsátó
háztartási kvarclámpákkal világítottuk
meg, képét pedig egy UV-érzékeny
videokamerával vettük föl, amelynek
objektívlencséje elé csak UV-t
áteresztõ szûrõt tettünk. Habár
a
napraforgó egész virágzata jól ismert
módon homogén sárga színû a
spektrum látható tartományában,
az UV-felvételen látható, hogy csak
a szirmok külsõ fele veri vissza az
UV-sugarakat. A szirmoknak a virágzat
középpontjához közelebbi fele,
valamint a virágzat belseje UV-elnyelõ.
Ilyen ultraibolya mintázat sok más
virágra is jellemzõ (a szerzõk felvétele) |
A virágok nagy része UV-elnyelõ háttér
elõtt nyílik (pl. levelek, talaj), ezért nekik elõnyös,
ha szirmaik UV-tükrözésével is rásegítenek
feltûnõ színeikre. Más virágok pontosan
az ellenkezõjére törekednek: pl. az UV-ben is nagy fényerõsségû
havas háttér elõtt a hóvirág éppen
az UV-elnyelõ szirmai révén látható
a beporzók számára. Ezen felül gyakoriak a növényi
UV-mintázatok is. Jellemzõ példa, amikor a számunkra
homogén színû szirmok belsõ része UV-elnyelõ,
külseje viszont UV-visszaverõ, ezzel jelezvén a nektárhoz
vezetõ utat (2. ábra). A méhek a szirmok szélét
pl. sárgának és egyben “UV-nek” is látják;
ezt a különleges színt nevezik méhbíbornak.
Egy fölöttébb érdekes táplálékkeresési
technikát alkalmaz a nálunk is honos vörös vércse.
A vándorló ragadozó madarakról, (mint amilyen
a vörös vércse is), általában ismert, hogy
az igen nagy, számukra ismeretlen, új területeket viszonylag
rövid idõ alatt fel tudják térképezni
a prédaállataik által sûrûn lakott területeket.
E gyorsaság rejtélyesnek tûnik. A vörös vércse
elsõsorban kisemlõsöket, pl. pockokat zsákmányol.
A pockok vizeletükkel és ürülékükkel
jelölik meg az ösvényeiket. E nyomok fényvisszaverése
340 nm-nél maximális, míg 370 nm-nél minimális,
azaz az utóbbi esetben erõsen elnyelik az UV-fényt.
Ily módon az UV-tartományban jelentõs kontrasztot
mutatnak a környezõ növényzethez képest.
Kiderült, hogy a vörös vércsék nem a szaglásuk
útján derítik fel a pockokat, ill. azok friss, vizelettel
és ürülékkel átitatott nyomait, hanem UV-látásuk
segítségével. A pocoknyomokat és az azok környezete
közti UV-kontrasztot észlelik. E madarak repülés
közben a mezei rágcsálók ösvényeit
figyelve UV-látásukkal gyorsan ki tudják választani
a nagy pocoksûrûségû területeket, ahol sokkal
eredményesebben vadászhatnak.
Fajon belüli és fajok közötti
UV-kommunikáció
Az UV-látás fajon belüli kommunikációra
is használható, fõként a párválasztásban.
A madarak bonyolult tollazata jó példa a többszörös
funkcióra: a hímek a színeikkel próbálják
elnyerni a nõstények tetszését és elriasztani
ellenfeleiket. Sok madárfaj tollazata visszaveri az UV-fényt
és feltûnõ mintázatot hordoz. Létezik
egy tollfajta, amely kimondottan alkalmas UV-minták létrehozására.
Ennek szivacsszerû szerkezete látszólag rendezetlenül
elhelyezkedõ keratinszemcsékbõl áll, amelyek
méretüknél fogva jól szórják az
UV-fényt. A rétegek közötti interferencia is szerepet
játszik, ugyanis az üregeket kitöltõ anyag törésmutatójának
változtatásával együtt módosul az UV-visszaverõképesség
is.
Az UV-mintáknak a vizuális kommunikációban
betöltött fontos szerepét bizonyítja pl. az is,
hogy a pekingi vörösbegyek elõnyben részesítik
azt a társukat, amelyet UV-fényt áteresztõ
plexiüvegen át láthatnak egy olyan madárral szemben,
amelyet UV-t elnyelõ üvegen át figyelhetnek. Noha a
színes testfelszín mutogatására általában
a madarakat említik példaként, az ibolyántúli
színek érzékelése a halaknál is kibõvítheti
a testükön látható jelek számát a
fajtársaik szemében (pl. a guppiknál).
A testfelszíni UV-mintázatokra további példát
a Puerto Ricoban élõ öt közelrokon gyíkfaj
szolgáltat. Mindegyik faj retinájában négy
látópigment (365, 450, 495 és 565 nm) található.
Kommunikációjukban fontos szerepet játszanak a vizuális
jelzések. Ennek egyik eszköze a színes, kifeszíthetõ
toroklebeny, amit majdnem kizárólag kommunikálásra
használnak; máskülönben elrejtik. Két gyíkfaj
toroklegyezõje sok UV-t ver vissza, egy harmadik fajé pedig
UV-mintázatot visel. A másik két faj toroklegyezõje
elnyeli az UV-fényt. Mind az öt gyíkfaj visel a szájzugában
egy-egy fényes UV-foltot is, amely csak akkor látható,
amikor kitátják a szájukat. E póz a gyíkok
jellemzõ fenyegetõ magatartása. A toroklebenyek eltérõ
UV-elnyelése korrelál az egyes fajok élõhelyének
optikai sajátságaival. Az elõbbi három faj
napfényes területen él, ahol a kék ég
is látható, tehát viszonylag erõs UV-sugárzás
éri õket. Ilyen körülmények között
az UV-fényt visszaverõ toroklebenyük fényes és
kontrasztos az UV-sugarakat alig visszaverõ háttérnövényzethez
képest. A másik két faj árnyas, erdõs
környezetben él, ahol kevés az UV-fény, így
kevés haszna lenne bármilyen UV-mintának a vizuális
kommunikációban. E tények azt sugallják, hogy
e gyíkok kommunikációjában fontos szerepet
játszik az UV-fény ott, ahol elég erõs ahhoz,
hogy megfelelõ kontrasztot biztosítson. Ezt a megállapítást
erõsíti az is, hogy az UV-mintákat csak toroklegyezõkön
találhatunk (ami speciálisan csak a kommunikáláskor
látszik), a test többi részén nem.
 |
 |
| 3. ábra. Különféle, preparált
lepkék felvétele látható (balra) és
ultraibolya (jobbra) fényben. (A) fecskefarkú lepke, (B)
kardoslepke, (C) sakktáblalepke, (D) citromlepke, (E) boglárkafaj.
A felvételek jól mutatják, hogy a lepkeszárnyak
látható fényben tapasztalható mintázata
sokszor jelentõsen eltér az UV-fényben érzékelhetõétõl.
Az “A” lepke elülsõ szárnyainak mintázata közti
eltérés azzal magyarázható, hogy a rovargyûjteményben
a lepke bal fölsõ szárnyáról már
kissé lekopott a hímpor (a szerzõk felvétele) |
A szárnyak és a test UV-színezete és mintázata
a lepkéknél is fontos szerepet játszhat mind a fajon
belüli, mind pedig a fajok közötti kommunikációban
(3. ábra). Segít elkerülni a hibridizációt,
ami bekövetkezhet, ha két közelrokon faj egyazon élõhelyet
népesít be. Sok fajban a látópigmentek spektruma
és az összetett szembeli eloszlása jól illeszkedik
a szárnyhoz és a visszavert fény spektrumához.
Így lehetõvé válik a faj hímjeinek megkülönböztetése
más fajok hasonló hímjeitõl. Egyes kutatók
adatai szerint viszont számos, azonos élõhelyen elõforduló
kéneslepke és citromlepke faj UV-szárnymintázatában
olyan nagy az egy fajon belüli variáció, hogy kétséges
a szerepük a fajmeghatározásban és a hibridizáció
megakadályozásában. Fõ szerepük inkább
a fajon belüli kommunikációban lehet. E lepkefajok hímjei
a szaporodáshoz csak a spermatofórájukkal járulnak
hozzá, amely hímivarsejteket és járulékos
mirigyeik váladékát tartalmazza (köztük
tápanyagokat is, amit a nõstény hasznosít).
A korábban már párosodott hímek ezen anyagokat
sokkal kisebb mennyiségben termelik, kimutathatóan csökkentve
ezzel a késõbbiekben velük párosodó nõstények
reproduktivitását és élethosszát az
olyan nõstényekhez képest, amelyek egy korábban
még nem párosodott hímet választottak. A párzás
ideje is megnõ és vele azonos mértékben a ragadozóknak
való kitettség veszélye. Tehát a nõsténynek
elõnyös, ha még nem párosodott hímet választ.
Ha elfogadjuk, hogy egy még nem párosodott hím
valószínûleg fiatal, akkor a nõsténynek
elõnyére válna, ha meg tudná állapítani
leendõ párja életkorát. Ez lehetséges
pl. a hímek UV-szárnymintázata alapján is.
E minták alapja a fényinterferencia, amely a szárnypikkelyek
mikroszkópikus lemezszerkezetén következik be. Ehhez
adódik a külsõ pikkelyek felületének bordázatán
létrejövõ interferencia, amely a helyzeténél
fogva az idõk során erõsen kopik és különbözõ
sérüléseket szenvedhet. Így a hímek UV-
szárnymintáinak állapota jelezheti tulajdonosuk életkorát
és ezzel a nõstények párválasztásának
alapja lehet.
Egy másik lehetõség, hogy a szárnymintázat
része a hímek egymás közötti párharcának.
A lepkék nõstényei általában egyedül
kóborolnak, míg a hímek gyakran találkoznak
egymással territóriumuk védelmében vagy a nõstényekért
vívott harcban. Eszerint a nagy távolságból
ható vizuális jelzések a hímek számára
fontosak, de a nõstények szemében kis jelentõségûek.
Ennek a hipotézisnek megfelelõen UV-mintákat rendszerint
csak hímekben találunk. Hat pillangófajban a hímek
és nõstények között a szárny által
visszavert fény polarizációjában is találtak
különbséget. Ennek is fontos szerepe lehet a fajon belüli
vizuális kommunikációban olyan fajok esetén,
amelyek szeme érzékeny a fénypolarizációra.
Ha egy, a szárnyain UV-mintázattal rendelkezõ rovar
összecsapja szárnyait, hirtelen eltûnnek az általuk
visszatükrözött UV-jelek is. Ez vagy repülés
közben, vagy a párzóhelyen történõ
szándékos szárnyrezegtetéskor észlelhetõ,
és felhívhatja a partnerek figyelmét. Ennek különösen
azokban a fajokban lehet jelentõsége, ahol az UV-receptorok
egyben kiváló rezgés-érzékenységgel
is bírnak.
Az UV-minták a lepkéknél a kommunikációban
fontosak, de az még vita tárgya, hogy a fajon belüli
vagy a fajok közötti kommunikációban használatosak-e.
Úgy tûnik, hogy a UV-mintázat számos egyéb
rovar- és gerinctelen faj kommunikációjában
is fontos szerephez jut.
A napi ritmus és az UV-fény
Az UV-fény napi ingadozása információt nyújt
az idõrõl és ezért felhasználható
a cirkadiális (huszonnégyórás) bioritmus kalibrálására.
Régóta ismert tény, hogy az UV-fénynek szerepe
van a gerinctelenek cirkadiális ritmusában, de újabban
kiderült, hogy fontos a gerincesek számára is. Például
mesterséges UV-megvilágítással átállítható
a kanárimadarak napi ritmusa. Az UV-fény képes szabályozni
a neuroendokrin és cirkadiális válaszokat néhány
rágcsálófajnál, pl. az aranyhörcsögnél
és a vándorpatkánynál. E rágcsálókban
az UV-megvilágítás blokkolja a reproduktív
rendszer rövid fotoperiódus-indukálta leállásait,
gátolja az éjszakai melatonin-szintézist, szabályozza
a melatonin-ritmust és eltolja az aktivitási ritmust.
Az írás az 1996. évi cikkpályázaton
I. díjban részesült.
Irodalom
[1] Goldsmith, T. H.: Hummingbirds see near ultraviolet light. Science
207, 786-788 (1980)
[2] Hárosi, F. I., Hashimoto, Y.: Ultraviolet visual pigment
in a vertebrate: a tetrachromatic cone system in the dace. Science 222,
1021-1023 (1983)
[3] Fleishman, L. J. et al.: Ultraviolet vision in lizards. Nature 365,
397 (1993) The Biology of Ultraviolet Reception. Vision Research (special
issue) 34, No. 11 (1994)
[4] Hollósy, F.: Az UV-B-sugárzás I., II. Természet
Világa 126, 258-262, 320-322 (1995)
[5] Tovée, M. J.: Ultra-violet photoreceptors in the animal kingdom:
their distribution and function. Trends in Ecology and Evolution 10, 455-460
(1995)
[6] Viitala, J. et al.: Attraction of kestrels to vole scent marks visible
in ultraviolet light. Nature 373, 425-427 (1995)
[7] Bennett, A. T. D. et al.: Ultraviolet vision and mate choice in
zebra finches. Nature 380, 433-435 (1996) Kísérõszöveg
az ábrákhoz
| Természet Világa, |
128. évf. 4. sz. 1997. április, 155 -159. o. |
|
https://www.kfki.hu/chemonet/TermVil/ |
|
https://www.ch.bme.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez