EVOLÚCIÓ
JORDÁN FERENC
Az evolúciós színjáték
fõszereplõi
A rókának fontos a nyúl, mert egyik kedvenc
tápláléka. A nyúlnak is fontos a róka,
mert tõle függ, hogy mennyire szaporodhat el. De vajon mennyire
fontos a nyúl és a róka az erdõ életében?
Minden faj egyenrangú vagy esetleg vannak kitüntetett szereplõk
az ökológia színpadán? Egyre inkább úgy
tûnik, hogy vannak. Ezek a kulcsfontosságú fajok, amelyek
egységes leírása és vizsgálata még
távoli célnak tûnik. A helyzetet nehezíti például,
hogy csak akkor derül ki, melyek a kulcsfajok, ha valamennyire az
egész életközösséget ismerjük már.
Az ökológiai munkák többsége viszont egy-két
kiemelt faj kapcsolatára összpontosít (pl. a ragadozó
és zsákmány viszonya). Szerencsére egyre több
az olyan átfogó jellegû kutatás, amely kezdi
megmutatni, melyek az evolúciós színjáték
fõszereplõi.
Néha akarva, néha akaratlanul, az ember új fajokat
telepít be egy-egy területre, más fajokat pedig kiirt,
vagy csak a populációikat pusztítja. A következmények
megfigyelése és az adatok összegzése arra a következtetésre
juttatta a kutatókat, hogy a természetben élõ
fajok nem azonos jelentõségûek az ökológiai
rendszerek mûködésében. Léteznek nagyobb
és kisebb szerepet játszó, szürkébb
fajok is. A kiemelkedõ fontosságú fajokat Robert Paine,
amerikai ökológus 1969-es szóhasználata alapján
kulcsfajoknak
(keystone species) nevezzük (az angolban a keystone a boltív
középsõ kövét jelenti, melynek hiányában
az egész boltív összeomlik).
Azóta a kulcsfajok egész garmadáját
leírták már, de mint ahogyan az ilyenkor lenni szokott,
a kezdeti lelkesedés zûrzavarhoz, tesztelhetetlen eredményekhez
és szubjektív következtetésekhez vezetett. Nyitott
a kérdés: mikor lesz egységes elmélet? Mikor
lehet a fajokat azonos szempontok alapján összevetni egymással?
Vajon érdemes-e általánosságokat keresni jellemzésük
során? A kulcsfajok szerepének tisztázása egyébként
természetvédelmi szempontból vagy a társulások
mûködésének mélyebb megértése
szempontjából is felbecsülhetetlen jelentõségû.
Kibõl lehet fõszereplõ? Szinte bárkibõl.
Kezdetben ugyan úgy tûnt, érdemes táplálkozási
kategóriák alapján szûkíteni a kört.
Paine elsõ ilyen témájú közleménye
egy olyan tengeri csillagról szólt (Pisaster),
amely a kaliforniai sziklás tengerpartok csúcsragadozója.
Azokon a területeken, ahol ez a csillag kipusztult (vagy eltávolították),
gyökeresen megváltozott az egész életközösség
viselkedése. Más fajok is eltûntek az érintett
területekrõl, majd újak jelentek meg és a megmaradó
fajok közötti kölcsönhatások mintázata
is átrendezõdött. Úgy tûnt tehát,
a Pisasternek közösségformáló szerepe
van. Ezt annak tulajdonították, hogy csúcsragadozóként
ez a faj a tengerpart ura, hatása közvetve vagy közvetlenül
mindenre kiterjed.
Ezzel egy idõben vált híressé az
Alaszkától egészen Dél-Kaliforniáig
elõforduló tengeri vidrák esete is. Az õslakók,
a telepesek és a természetkedvelõ sportolók
hatékonyan ritkították ezeket az állatokat,
egyes helyeken ki is irtották õket. Ahol kipusztultak, elszaporodott
kedvenc csemegéjük, a tengeri sün. A sok sün
pedig nagy étvággyal fogyasztotta a part menti moszaterdõket,
ahonnan a kisebb halak és más lakóik is kipusztultak.
Így a vidrák kiirtásának hatására
sünsivatagok keletkeztek (1. ábra). A vidrák
tehát bár ez azonnal nem szembetûnõ komoly
szerepet töltenek be a part menti vizek életében. Ahol
sikerült a vidrákat visszatelepíteni, az eredetihez
nagyon hasonló közösség népesítette
be ismét a partot, a sünsivatagok pedig megszûntek.

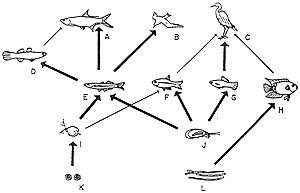
1. ábra. A kaliforniai tengerpartok ura a tengeri vidra.
Ha kipusztul, gyors szaporodásnak
indul legfontosabb tápláléka, a tengeri sün.
Elõbb-utóbb viszont ez a moszat- és
hínártelepek eltûnéséhez vezet,
melynek következtében a dús növényzet többi
lakója is eltûnik. Végül kialakulnak a
sünsivatagok (Bond, 1994 nyomán)
A tengeri csillagok és a vidrák példája
nyomán szinte azonnal elterjedt az a téveszme, hogy kulcsfaj
csak csúcsragadozó lehet. Megszületett az a felismerés
is, hogy a kulcsfajok lényegében kijelölik az egyes
életközösségek határát is azáltal,
hogy hatásuk kaszkádszerûen kiterjed mindenre, ami
a táplálékhálózatban alattuk helyezkedik
el. Ebben lehet is némi igazság, ha a kulcsfaj valóban
csúcsragadozó. A nyolcvanas években azonban azt is
megfigyelték, hogy a táplálékhálózat
közepe táján elhelyezkedõ fajok is meghatározók
lehetnek. Sokat emlegetett példa erre a kengurupatkány, amely
gödröket és üregeket ás, így különösen
a kisebb állatok és a növények környezetét
képes alapvetõen megváltoztatni. Kimutatták,
mekkora hatással van ez a tevékenység a teljes vegetációra
és azon keresztül természetesen az állatvilágra
is. A kengurupatkányok eltûnése vagy felbukkanása
képes tehát átformálni a társulások
fajösszetételét. Az eddigiekkel szemben fontos különbség,
hogy itt már nem táplálkozási kapcsolatokon
keresztül érvényesül a kulcsfaj hatása.
A kilencvenes években többen rámutattak arra
is, hogy esetenként a termelõk is lehetnek kulcsfajok, bár
valamilyen okból ennek elismerésétõl kezdetben
húzódozott a szakma. Nagy szerepet játszanak például
egyes fügefélék a trópusokon, vagy a talajlakó
kékbaktériumok és zuzmók a Negev-sivatagban.
A kulcsfajnak bizonyuló termelõ szervezetek trofikus hatása
a táplálékhálózatban felfelé
terjed, ellentétes irányban, mint a csúcsragadozóké.
Emellett akár egyszerûen csak mennyiségük miatt
is fõszereplõvé válhatnak. Például
egy tölgyesben a tölgy vitathatatlanul kulcsfaj, meghatározó
a közösség összetételében és
kipusztulása minden más élõlényre tragikus
hatással lenne.
Újabban kulcsfajként emlegetnek lebontó
élõlényeket is, már amennyiben az esetükben
gyakran homályos fajfogalom ezt megengedi. A kulcsvírusok
és kulcsparaziták is kezdenek divatba jönni. A paraziták
mellett keresik a gazda típusú kulcsfajokat is (például
néhány pálmafajt már kulcsgazdának tartanak).
Kulcskritériumok
Láttuk, hogy egyik fõ táplálkozási
csoportból sem lehet kizárni a kulcsfajokat. Ez a megközelítés
tehát nem vezetett túl sok eredményhez, de szerencsére
más szempontok alapján is igyekeztek osztályozni ezeket
a fajokat. Az egyes próbálkozások során mindenki
megalkotta a saját kulcsfaj-definícióját, de
elfogadott meghatározás híján eddig elég
szubjektív és plasztikus maradt ez a szakkifejezés.
E. O. Wilson kicsit talán komolytalan kulcsfajfogalma
a következõ: a kulcsfajok a nagy játékosok
(big players). Ez a meghatározás ugyan valóban nem
túl hasznos, de remekül tükrözi az általános
szemléletet és hozzáállást, amely gyakran
szubjektív és anekdotikus. Nem sokkal kreatívabb az
(egyik) Ecology címû tekintélyes szakkönyv meghatározása
sem, amely szerint kulcsfajok azok, amelyek különösen alapvetõ
és nagy hatású kölcsönhatásokat alakítanak
ki fajokkal.
Valamivel konkrétabb Paine legelsõ megfogalmazása,
mely szerint kulcsfaj az, ami meghatározza a társulás
integritását és idõben változatlan fennmaradását.
Ezen a definíción kicsit érzõdik, hogy megalkotásakor
csak a csúcsragadozókra gondoltak. Ennek ellenére
jól sejteti a kutatások egyik fõ célját:
valamilyen kapcsolatot találni a társulás stabilitása
és egy-egy faj sorsdöntõ jelentõsége között.
Mindenesetre a tengeri csillag és a vidra esetéhez jól
passzol.
Sokszor egyszerûen azt tekintik kulcsfajnak, ami gyakori.
Lehet persze összefüggés, mondjuk egy erdõalkotó
fa egyben kulcsfaj is, de felesleges lenne új fogalmat bevezetni,
ha pusztán csak errõl lenne szó.
1987-ben Jackson és Kaufmann beszámolt
egy ellenpéldáról. A Diademaantillarum nevû
tengeri
sün a Karib-térségben él, csúcsragadozó
és rendkívül gyakori. Automatikusan kulcsfajnak tartotta
mindenki, amíg egy-egy területrõl ki nem pusztult. Ekkor
viszont feltûnõen nem történt semmi. Az ok még
nem tisztázott, a többi faj lárváinak tulajdonságaival,
illetve a fogyasztott növények populációdinamikájával
lehet kapcsolatos a sünök csendes távozása. Mindenesetre
jól mutatja, hogy a gyakoriság nem jelent feltétlenül
fõszerepet a közösség életében. Persze
az sem zárható ki, hogy a sünpusztulás hatása
csak lassan bontakozik majd ki.
1994 decemberére konferenciát hívtak össze
a Hawaii-szigeteken a kulcsfajok helyzetének tisztázására.
A tizenöt neves résztvevõ természetesen a definíciók
egységesítésével is foglalkozott. A végsõ
verzió szerint kulcsfaj az, amelynek hatása a közösség
egyéb tagjaira nagy, legalábbis nagyobb, mint amit az adott
faj tömegessége alapján várnánk. Ez nagyjából
David Tilman ötletének felel meg, aki szerint valamilyen
módon egy hatás/biomassza arány alapján kellene
a fajokat rangsorolni jelentõségük szempontjából.
Kicsit más jellegû Stuart Pimm és
Michael Gilpin ötlete. Szerintük kulcsfaj az, aminek a
kihalása a legtöbb másodlagos kihalást okozza.
A másodlagos kihalás egy elsõdleges kihalási
eseményt gyorsan követ és egyértelmûen
annak hatására vezethetõ vissza. A legtöbb példa
esetében ez a meghatározás logikusnak tûnik,
és hasznos lehet mint a további vizsgálódások
kiindulópontja. Természetesen kihalás alatt itt szerencsére
legtöbbször csak valamilyen lokális kipusztulásra
kell gondolni, azaz csupán egy-egy populáció eltûnésére.
Clive Jones és társai szenvedélyesen
kutatják azokat a fajokat, amelyek nem táplálkozási
kapcsolataik révén fejtenek ki jelentõs hatást
a társulás más tagjaira, hanem valamilyen módon
jelentõsen átalakítják az élettelen
környezetet és így befolyásolhatják
sok más faj viselkedését is. A fentebb bemutatott
kengurupatkány is közéjük tartozik, valamint például
a levélvágó hangyák, a hódok vagy a
kisebb köveket hatalmas mennyiségben felaprózó
sivatagi csigák. Ezek a fajok mind olyan mértékû
átalakítást végeznek környezetükön,
mely sok más faj számára meghatározó.
Végül a kulcsfajok közé tartozhat egy
nagyon fontos, mutualista, kölcsönösen elõnyös
kapcsolatokat kialakító faj is. Ilyen például
az a sok növény beporzásában fontos kolibri,
amelynek eltûnése több növény szexuális
életét keserítené meg, s ezzel hatással
lenne a társulás majd minden tagjára. Bár más
szempontok alapján talán fontosabbnak tûnnek a szélsõségesen
specializálódott mutualisták, de lokális hatásuk
révén mégis jóval kisebb szerepet játszanak
az életközösség életében.
Gyanús idegenek
Az eddigiekben fajok kihalásáról vagy lokális
eltûnésérõl volt szó. Sokszor a jövevények
is fõszerephez juthatnak. Hatásuk gyakran tragikus, fõleg
ha mesterséges betelepítések alanyai. A sok példa
közül talán a legismertebbek az ausztráliai üregi
nyulak, de egzotikusabb egy trópusi hal esete.
A Cichla ocellaris egyike az Amazonas medencéjében
élõ rengeteg halnak. A horgászok szeretik és
ráadásul még szép is. Elõnyös tulajdonságai
miatt egy üzletember 1965-ben betelepítette egy panamai elszigetelt
tavacskába. A cél az volt, hogy a cég alkalmazottainak
felhõtlen kikapcsolódási lehetõséget
szerezzen. A hirtelen jött nagy esõzések hatására
azonban a tavacska elszigeteltsége megszûnt és közös
víztér alakult ki a Rio Gatuncillo nevû kis patakkal,
mely a Gatun-tavat tápláló Chagres-folyóba
ömlik. A halak kihasználták a kínálkozó
alkalmat, és kipróbálták az életet a
kis patakban. 1967-ben megjelentek a Chagresben, 1969-ben pedig a Gatun-tó
peremén. 1972-ben szinte az egész tavat benépesítették
már, azaz kb. 5060 kilométert tettek meg hét év
alatt. Mindez nem is lett volna nagy baj, ha ezzel csak a horgászok
lehetõségei bõvülnek. A Gatun-tó élõvilága
azonban érzékenyen reagált a jövevényre
(2.
ábra). A többi hal közül szinte alig volt olyan,
amelyiket ne zavarta volna meg a Cichla jelenléte. Sok teljesen
eltûnt a tóból, mások viszont eddig nem látott
mértékben elszaporodtak. A közvetett hatások
hasonlóan drasztikusak voltak, a zooplankton és a rovarok
esetében éppen úgy, mint a patak gerinces faunájának
életében. Összességében a másodlagos
lokális kihalások sokkal jellemzõbbek voltak, mint
az egyedszám növekedése egy-egy populációban.
A táplálékhálózat és a társulást
alkotó fajok kapcsolatrendszere gyökeresen megváltozott.
A helyi horgászokat leginkább talán az bosszanthatta
fel, hogy a rovarevõ halak csökkenésével elszaporodtak
a szúnyogok (pont azok, amelyek egyébként a malária
kórokozóját is terjesztik). A Gatun-tóban a
Cichlaocellaris
egyértelmûen kulcsfajjá lépett elõ, alapvetõen
meghatározva a közösség összetételét
és a fajok kapcsolatrendszerét.
 |
 |
| a) |
b) |
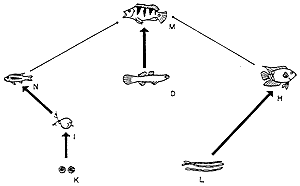
2. ábra. A panamai Gatun-tó
vázlatos táplálékhálózata a Cichla
ocellaris halfaj behurcolása elõtt
(a) és azt követõen (b). A közösség
összetétele némileg leegyszerûsödött,
sok faj nem tudott alkal-
mazkodni a jövevényhez. A Cichla egyedül
csúcsragadozóvá lépett elõ (M-mel jelölve),
többek
között saját fajának fiataljait fogyasztva
(N jelöli a fiatal Cichlát). Sok más halfaj (F, G) és
néhány
madár (B, C) ugyanúgy eltûnt a tó környékérõl,
mint egyes rovarok (J). Az új halfaj erõsen befolyá-
solta a teljes közösség viselkedését.
Más betelepítéseknél a hatás jóval
erõsebb is lehet, habár szinte
mindig kedvezõtlen (Zaret és Paine, 1973 nyomán). |
Kulcstulajdonságok
A példák alapján most már megpróbálhatjuk
összegezni, milyen is egy tipikus kulcsfaj. Táplálkozási
szempontból bárhova tartozhat, lehet ragadozó vagy
akár növény is. Valószínûleg a társulás
típusától sem függ a kulcsfajok jelenléte,
azaz a Föld minden vidékén elõfordulhatnak az
átlagosnál nagyobb hatású fajok.
Fontos felismerés, hogy e kulcsszerep erõsen függhet
a körülményektõl. A Pisaster tengeri csillag
például az újabb kutatások alapján csak
a hullámverte sziklákon tekinthetõ kulcsfajnak, a
védettebb területeken jelentõsége sokkal kisebb.
A Gatun-tó rettegett jövevénye pedig távolról
sem volt fõszereplõ az Amazonas környékén.
A kulcsfajok jelentõsége függ a diverzitás
aktuális mértékétõl is. Ha a diverzitás
csökken, a megmaradó fajok jelentõsége növekszik.
Többször felvetõdött már, vajon mit mondhatunk
a kulcsfajok és a többi faj közti kapcsolatok erõsségérõl.
Úgy tûnik, általában kevés fajt tarthatunk
csak kulcsfontosságúnak egy adott területen. Ezek viszont
vagy szinte mindenki mással közvetlen kapcsolatban vannak,
vagy csak kevesebb, de nagyon erõs kapcsolatot alakítanak
ki másokkal.
Sokszor nemcsak az egyes fajok, hanem néhány faj
szorosan összetartozó csoportja is fõszerepet játszik
a társulás életében. Az ilyen kulcsfajkomplexek
jelentõségét csak mostanában kezdik hangsúlyozni.
Szép példát mutatott be erre néhány
éve Daily és két társa. Egy coloradói
hegyi erdõben azt vizsgálták, vajon melyek a meghatározó
fajok (3. ábra). Az eredmény: egy harkály,
két fa és egy gomba. A harkály egyik fontos tevékenysége,
hogy olyan üregeket váj a gombával fertõzött
nyárfákba, melyeket más madarak is használnak.
Másik foglalatosságaként megcsapolja a fûzfák
nedveit, melybõl rajta kívül kolibrik, mókusok
és más állatok is táplálkoznak. Úgy
tûnik, azokban az erdõkben, ahol a fenti négy fõszereplõ
mindegyike jelen van, nagyon hasonló a közösség
összetétele. A négy fajból álló
kulcsfajkomplexre mintegy ráépül mindenhol ugyanaz a
fajgarnitúra. Ahol a négy fajból akár csak
egy is hiányzik, a társulás szerkezete másképpen
formálódik. Az ilyen erdõk egymástól
is sok szempontból különböznek. A harkály,
a két fa és a gomba jelenléte alapján tehát
következtethetünk nagyjából a társulás
többi tagjára is. Bizonyos szempontból persze egyenként
is kulcsfajoknak tekinthetõk, mégis érdemes õket
kulcsfajkomplexként együtt tárgyalni, legalábbis
a társulás szervezõdése szempontjából,
mert így érdekesek.
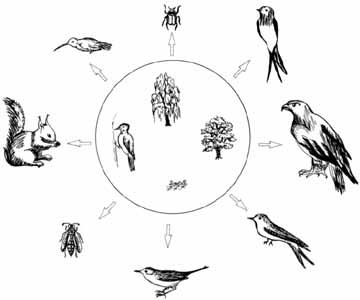
3. ábra. Egy harkályfaj, egy fûz- és egy
nyárfafaj, valamint egy farontó gomba alkotja
a coloradói hegyekben az erdei társulások
gerincét. Ha õk négyen megélnek együtt
egy területen, sok más faj jelenlétére
is következtetni lehet. A négy fajból álló
kulcs-
fajkomplex mûködésérõl bõvebben
a szövegben (Daily és társai, 1993 nyomán)
A kulcsfajok szinte mindegyik meghatározása tartalmazza
valamilyen formában a fajok hatását, vagy a köztük
lévõ kapcsolatok erõsségét. Ezek viszont
egyrészt nehezen mérhetõ mennyiségek, másrészt
csak akkor tudunk rájuk következtetni, ha már kihalt
a vizsgálni kívánt faj és ennek a sajnálatos
eseménynek a hatásait elemezzük. A természetvédõknek
persze nem okoz túl nagy boldogságot, ha felmérhetik
egy éppen kihalt faj jelentõségét. Sokan rágódnak
azon, vajon hogyan lehetne a kulcsfajokat elõre megmutatni.
Amíg ki nem pusztul egy faj, és nem látjuk ennek hatását,
általában csak tippelni tudunk, mennyire is fontos az másoknak.
Egyelõre sajnos semmilyen lehetõség nincs
arra, hogy számokkal jellemezzük a kulcsfajokat. Az egyetlen
eddigi ismert próbálkozás egy olyan index (CI, community
importance) kidolgozása, amely a fajok jelentõségét
méri. A szempont a másodlagos kihalások mértéke,
azaz az adott faj kihalását követõ és
arra visszavezethetõ kihalásoknak a teljes fajszámhoz
viszonyított aránya. Persze itt is csak utólag lehet
okosabb a kutató és legfeljebb összehasonlító
jellegû következtetéseket vonhat le más társulásokra.
A CI-indexet nem is nagyon használják semmire, legfeljebb
megemlítik, mert jól mutat mint a kulcsfajokra vonatkozó
egyetlen matematikai jellegû kifejezés. A megfelelõ
kvantitatív háttér kidolgozása lehetõvé
tenné, hogy ne csak két kategóriába sorolhassuk
a fajokat, úgymint kulcs és normális. Minden fajhoz
érdemes lenne hozzárendelni valamilyen mennyiséget,
mellyel jelentõségét mérhetnénk a CI-indexhez
hasonlóan, de annál valamivel hatékonyabban. Átmenetileg
ezt az interakciós erõsségek becslésével
próbálják pótolni, de sok lehetõség
van még a továbblépésre. Ilyen elemzések
segítségével becsülni lehetne például:
vajon mennyiben tekinthetjük minõségileg is másoknak
a kulcsfajokat?
Néhány esetben mesterséges beavatkozások
hatását vizsgálják, nem várnak a fajkompozíció
természetes megváltozására. A behurcolt fajok
példái is tekinthetõk ilyen kísérleteknek,
ha gyakran nem is szándékosak. A vizsgálatok egyik
hátránya az, hogy sokszor nagyon lassan reagál a társulás
a zavarásokra. A reakcióidõ kisebb a tengerekben,
emiatt a legtöbb használható eredmény a vízi
közösségekre vonatkozik.
De vajon érdemes-e a kulcsfajok viselkedésének
általános törvényszerûségeit keresnünk?
Vajon a mérhetetlen bonyolultság nem mossa-e el az amúgy
jellegzetes tulajdonságokat? Lehet, hogy a Gatun-tótól
pár száz kilométerre fekvõ másik tóban
a Cichla egyáltalán nem terjedne el, talán
rögtön ki is pusztulna. Nem tudjuk még megmondani, mekkora
az egyes esettanulmányok tanulságainak érvényességi
köre.
Szürke mellékszereplõk
Talán sokakban felvetõdik a kérdés, nem
tekinthetõ-e végsõ soron minden faj kulcsfajnak, hiszen
minden kihalásnak vannak következményei, minden faj
fontos valamelyik másiknak. Érdemes-e a kisebb-nagyobb különbségek
miatt egyeseket kiemelni? Érdemes. Léteznek ugyanis olyan
fajok, amelyek ökológiailag szinte egyenértékûek
másokkal. Lényegében azonos niche-t töltenek
be és tulajdonképpen felcserélhetõk (redundánsak).
Ezek a fajok bizonyos szempontból már valóban alapvetõen
eltérnek a kulcsfajoktól. Elsõsorban persze az alacsonyabb
rendûek között ismertek ilyen példák, tehát
az algák, a baktériumok vagy mondjuk a zooplankton fajainál.
Sok vita folyik arról, mekkora lehet a feleslegesnek látszó
fajok aránya az ilyen esetekben. A redundancia hívei szerint
egyes fajok eltûnhetnek anélkül, hogy bárki észrevenné
azt. A helyzetet színesíti, hogy a közös funkcionális
csoportot alkotó fajok közül minden évben más
fordul elõ tömegesen és a többi csak minimális
egyedszámban, aztán a következõ évben
fordul a kocka. Egy karibi szigeten dolgozó kutatók például
azt figyelték meg, hogy az algaszõnyeg egy adott helyen mindig
nagyjából ugyanúgy néz ki, de két egymást
követõ évben szinte egy azonos fajt sem találtak.
Aztán hoszszabb távon megmutatkozott a vetésforgó
jelleg. Valószínû, hogy mondjuk egy 78 fajból
álló funkcionális csoport egyik tagjának kipusztulása
semmilyen észrevehetõ változást nem okozna,
de ennek ellenére a redundancia hívei is óva intenek
attól, hogy az ilyen fajoknak kevesebb figyelmet szenteljünk
(például a természetvédelem területén).
Egy jó ökológus sosem lehet elég biztos a dolgában.
Ugyanilyen funkcionális csoportokról beszélnek
a baktériumok esetében is. Itt inkább azért,
mert a fajfogalom kényesebb és sokszor nehezen értelmezhetõ.
A Rhizobiumokról például tudják, hogy nitrogént
kötnek, de nem érdemes feszegetni, hogy pontosan mely fajaik
vannak jelen egy adott társulásban. A lényeg: ha vannak
Rhizobiumok, akkor van nitrogénkötés is. Ennek jelentõsége
meghatározó a társulás mûködése
szempontjából, de sokszor meg sem próbálnak
kulcsfajokról beszélni.
A funkcionális csoportok és a kulcsfajkomplexek
közti különbséget talán érdemes hangsúlyozni.
Elõbbiben mindegyik képes elvégezni egy adott funkciót,
de nem feltétlenül egyszerre. Utóbbiban viszont mindegyiknek
megvan a maga szerepe, és azt egyszerre végzik el.
A kulcsfajokról szerzett ismeretek alátámasztják
azokat az elképzeléseket, amelyek szerint pusztán
a fajlisták hosszának tanulmányozása valószínûleg
nem vezet helyes természetvédelmi stratégiákhoz.
Az életközösségek mûködésének
vizsgálata más preferenciákat indokolhat. Ha például
nincs pénz mindenkit megvédeni, nem mindegy, kit védünk.
A kulcsfajok védelme pedig hatékony, mert segítenek
fenntartani az ökoszisztémák integritását.
Más megfogalmazásban: elvesztésük lavinaszerû
kihalási folyamatokat indíthat el. A fajok kipusztulása
pedig az egyre súlyosabb globális problémák
alapja. A környezet kihalásokat okozó hirtelen megváltozásai
(pl. katasztrófák) mellett egyre jobban ismerjük már
a biológiai rendszerek olyan belsõ tulajdonságait
is, amelyek szintén hozzájárulhatnak a fajok számának
csökkenéséhez. A fajok közötti bonyolult kölcsönhatások
eredményeként gyakran amúgy jól alkalmazkodott
fajokat is megcsaphat a kihalás veszélyének fuvallata.
Ha alaposabban megismerjük az egyes fajok jelentõségét,
pontosabb képet kaphatunk a veszélyeztetett fajokról
is, és jobban megérthetjük egy-egy faj hosszabb távú
viselkedését. Ez pedig hasznos eszköze lehet a természetvédelemnek.
A kulcsfajok feltérképezése mindenféle
alkalmazástól függetlenül is izgalmas feladat,
akár az elméleti biológus számára is.
A hozzá nem értõ pedig remélhetõleg
sosem fogja az ilyen jellegû ismereteket a valódi fajüldözés
alapjául felhasználni.
Cikkpályázatunkon a szerzõ második díjban
részesült.
IRODALOM
Bond, W. J. 1994 Keystone species. In: Schulze, E. D. and Mooney, H.
A. (eds.): Biodiversity and Ecosystem Function, Springer Verlag, Germany.
Daily, G. C. et al. 1993 Double keystone bird in a keystone
species complex. PNAS, 90: 592-594.
Jackson, J. B. C. & Kaufmann, K. W 1987 Diadema antillarum was
not a keystone predator in cryptic reef environments. Science, 235: 687-689.
Zaret, T M. & Paine, R. T 1973 Species introduction in a tropical
lake. Science, 182:449-455.
| Természet Világa, |
130. évf. 2. sz. 1999. február, 6972. o.
https://www.kfki.hu/chemonet/TermVil/
https://www.ch.bme.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez