A szétszóródás evolúciója
Az állatok többsége élete során
folytonosan változtatja a helyét, szülõhelye
ritkán egyezik meg szaporodó- és táplálkozóhelyével.
Közismert, hogy az emlõsöknél a hímek fiatalon
elhagyják a szülõhelyüket, és távolabb
keresnek párt maguknak. A madaraknál általában
éppen fordítva van, a fiatal nõstények a kalandvágyóbbak.
Hasonló jelenség figyelhetõ meg a helyhez kötött
növényeknél is. Mivel maguk mozogni nem tudnak, magjaikat
szórják szét, igen sokféle technikát
alkalmazva. Utazhat a mag a szél hátán, egy madár
tollába akadva, vagy egy rágcsáló gyomrában
is. Természetesen adódik tehát a kérdés:
miért változtatják a helyüket az élõlények,
milyen szelekciós hatások miatt vált a szétszóródás
a legtöbb élõlény életének részévé?
Az evolúcióbiológia alapfeltevése, hogy az élõlények viselkedése, testfelépítése, egész életmenete, az adott körülményeket és kényszerfeltételeket figyelembe véve optimális vagy ahhoz igen közeli állapotú. Esetünkben az optimális azt jelenti, hogy az adott tulajdonság a lehetõ legnagyobb rátermettséget (azaz a legtöbb ivarérett utódot) biztosítja a tulajdonság hordozóinak. Tehát abból indulunk ki, hogy a szétszóródás, diszperzió általában növeli a rátermettséget, ezért olyan elterjedt tulajdonság.
Az elkövetkezendõkben a diszperziót támogató legfontosabb mechanizmusokkal ismerkedünk meg. Elõször azokat az eseteket vesszük sorra, amikor az ún. feltétlen szétszóródás is hatékonyan növeli a rátermettséget. Ezután megmutatjuk, hogy a környezethez érzékenyebben alkalmazkodó, feltételes diszperziós stratégiák magyarázatul szolgálhatnak további terjedési jelenségekre. Végül röviden összegezzük a fõbb megállapításokat, és megemlítjük a nehézségeket is.
Mikor elõnyös a szétszóródás?
Feltétlen diszperzió
Ha a szétszóródás a külsõ körülményektõl és az egyedek biológiai tulajdonságaitól függetlenül történik, akkor feltétlen diszperzióról beszélünk. Bár a valóságban az egyedek terjedése soha sem tekinthetõ teljesen feltétel nélkülinek, a magvak diszperziója vagy számos ízeltlábú terjedése megközelítõen ilyen. Feltétlen diszperziót feltételezni elsõsorban a probléma elméleti tárgyalásának megkönnyítése miatt érdemes. Ha egy egyszerûbb diszperziós stratégia is elõnyös, akkor egy kifinomultabb stratégia még inkább fokozza a rátermettséget.
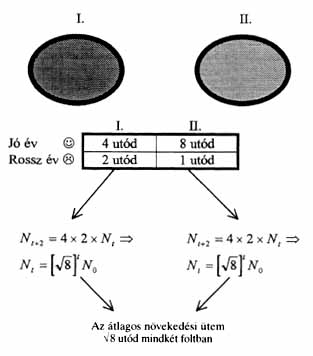
1.a ábra. A térben és idôben változó
élôhely modellje. Az élôhely két foltból
áll, ahol a "jó" és a "rossz" évek véletlenszerûen
váltogatják egymást. Nt-vel a foltban lévô
egyedek számát jelöltük a t-ik évben. A
t+2-ik évben 8Nt egyed lesz, ezért a populációk
átlagos éves növekedési üteme Ö8
mindkét foltban. (N0 a populáció mérete
kezdetben.)
Az a környezet, melyben az élõlények térben és idõben élnek, folytonosan változik. Ezért erõs lehet a gyanú bennünk, hogy a tér- és idõbeli heterogenitás jobban kiaknázható, ha az egyedek mozognak az élõhelyükön. Készítsünk egy nagyon egyszerû modellt. Tételezzük fel, hogy képzeletbeli populációnk egyedeinek élõhelye két foltból áll. Az egyedek minden évben egyszer szaporodnak, majd ezt követõen elpusztulnak. Az egyszerûség kedvéért tételezzük fel továbbá, hogy vizsgált populációnk szempontjából jó, illetve rossz évek vannak. Elég hosszú idõszakot vizsgálva a jó és a rossz évek fele-fele arányban jelentkeznek. Esetünkben az I. foltban jó évben átlagosan 4 utódot lehet felnevelni, a rossz évben 2-t. Ezzel szemben a II. foltban a jó év 8 utódot hoz, míg rossz évben csak 1 utódra van elegendõ táplálék (1.a ábra). Ez például úgy képzelhetõ el, hogy az I. foltban kiegyenlítettebb, míg a II. foltban szélsõségesebb mikroklíma uralkodik. Ahogy az 1.a ábrán bemutatott kis számolás alapján látható, a populáció átlagos növekedési üteme mindkét foltban azonos. Tehát az ivarérett felnõttek rátermettsége mindkét foltban megegyezik. Úgy tûnik, semmi elõnye nem származna egy mutáns típusnak abból, ha utódainak egy része (függetlenül a folt típusától és az adott év minõségétõl) áttelepülne a másik foltba. De ne bízzunk annyira a megérzéseinkben, inkább alakítsuk át a modellt úgy, hogy az utódok terjesztését megoldó mutáns rátermettségét is ki tudjuk számolni! A számolás egyszerûsítése érdekében tételezzük fel, hogy a mutáns típus utódainak a fele még születése évében átkerül a másik foltba. Bármelyik élõhelyen jelenik is meg a mutáns, utódainak fele átkerül a másik foltba. Tehát a második évben már ugyanannyi mutáns lesz mindkét élõhelyen. Ha ekkor éppen egy jó év következik, akkor az I. foltból 2 utód kerül át a II. foltba, a II-ból viszont 4 az I-be. Végül 6 mutáns utód lesz mindkét foltban. Hasonló okok miatt a rossz évben átlagosan 1,5 mutáns utód lesz mindkét foltban (1.b ábra). (Az 1,5 utód úgy értendõ, hogy a rossz évek egyik felében 2, a másik felében 1 utód lesz egy adott foltban.) Számításaink szerint tehát a diszperziót elõidézõ mutáns növekedési üteme átlagosan 3 utód (1.b ábra), ami fölülmúlja a helyben szaporodó típus növekedési rátáját (Ö8). Tehát egyre nagyobb arányban lesz a mutáns a populációban, végül teljesen kiszorítja a helyben szaporodót. Mivel a lényegesen összetettebb modellek vizsgálatai is erre az eredményre vezetnek, megállapíthatjuk, hogy ha az adott populáció élõhelye térben és idõben heterogén, akkor a lehetõ legkönnyebben megvalósítható, feltétlen diszperzió evolúciósan támogatott.

1.b ábra. A térben és idôben változó
élôhely modellje, ha diszperzió is van. Az élôhely
két foltból áll, ahol a "jó" és "rossz"
évek véletlenszerûen váltogatják egymást.
A mutáns az utódok felét a szomszéd foltba
helyezi. Nt-vel a foltban tévô egyedek számát
jelöltük a t-ik évben. A t+2-ik évben az 1. ábra
számolása alapján 9Nt egyed lesz, ezért
a populációk átlagos éves növekedési
üteme Ö9=3 mindkét foltban.
Természetesen a diszperzió sokszor veszélyes: a szétszórt magok többsége terméketlen talajba hull, az elvándorló pókcsemeték ragadozók áldozatává válnak stb. Kimutatható, ha a diszperzió veszélyeit is figyelembe vesszük, akkor is érdemes terjeszteni az utódokat, ha az elvándorló utódok túlélési esélye nagyobb egy kritikus értéknél, amit a foltok tér- és idõbeli heterogenitása határoz meg. Minél nagyobb a heterogenítás és minél kisebb a kapcsolat az egyes évek és helyek között, annál kisebb túlélési esély is elegendõ a diszperzió fenntartásához.
Az élõhelyfoltok idõbeli változékonyságának említésekor legtöbbször külsõ idõjárásbeli változásokra gondolunk, de ezt a változást okozhatja a populációk (vagy élõközösségek) belsõ dinamikai tulajdonsága is. Régóta ismert, hogy a populáció-dinamikai modellek bizonyos esetekben kaotikusan viselkednek, azaz bár a szaporodást leíró egyenletek determinisztikusak, a populáció viselkedése kiszámíthatatlan, véletlenszerû lesz (lásd a szerzõ írását folyóiratunk 1998 augusztusi számában a szerk.). Tehát, ha a populáció kaotikus, és az élõhely idõben nem heterogén, akkor egyedül a térbeli heterogenitás is elegendõ a diszperzió szelekciójához.
A terepmegfigyelések alapján azonban úgy tûnik, hogy a természetes populációk ritkán kaotikusak. Ezért, ha a környezet térben és idõben közel állandó, akkor nem várhatjuk, hogy a diszperzió elõnyös viselkedés lesz. Ahogy a következõ modell segítségével bemutatom, erre a megérzésünkre sem érdemes hagyatkozni.
Képzeljünk el egy szûznemzéssel szaporodó
populációt, ahol a nõstény egy kiválasztott
petézõhelyre több (esetünkben 4) petét is
rak. A peterakást követõen a nõstény elpusztul.
A kikelõ peték erõs versenyben állnak egymással,
ezért csak a legéletképesebb fogja megérni
a felnõttkort. Tehát a nõstény helyét
a következõ évben egyik utódja fogja átvenni.
A környezet változatlan, de az utódok között
a versengés igen erõs. Tegyük fel, hogy megjelenik egy
mutáns nõstény, amely a négy petébõl
hármat a környezõ petézõhelyeken rak le.
Esetleg a mutáns utódok közül fog három
átrepülni a szomszéd élõhelyek egyikére.
Ha evvel a stratégiával átlagosan egynél több
utódot tud a mutáns a következõ generációba
juttatni, akkor elterjed a populációban. Vizsgáljuk
meg tehát, hány utóda lesz az elterjedt típusnak,
és hány a mutánsnak. Azokban a foltokban, ahova a
mutáns utóda is eljut, az elterjedt típusnak átlagosan
4/5 utódja lesz, ahova a mutáns nem jut el, ott változatlanul
egy utódot hagy maga után, míg a mutánsnak
1+3·1/5 = 1,6 utóda lesz (2.a ábra). Megéri
tehát a szülõnek távolabbi petézõhelyeket
is fölkeresni, illetve érdemes a kikelõ utódoknak
elvándorolni a környezõ élõhelyekre.
 |
 |
| 2.a ábra. A diszperzió elônye állandó környezetben, ha az utódok között erôs versengés van. A modellben a szûznemzéssel szaporodó nõstény négy petét rak minden petézõ helyre (fehér kör), de ezek közül csak egy éri meg a felnôttkort. A mutáns típus a négy petébôl hármat a szomszédos levélre rak (fekete kör). | 2.b ábra. A diszperzió elônye állandó környezetben, ha az utódok között erôs versengés van. A modell ugyanúgy épül föl, ahogy a 2.a ábrán ismertetett modell. A különbség csupán annyi, hogy a három szétszórt petébôl átlagosan csak egy marad életképes. A sikertelen diszperziót szaggatott vonallal jeleztük. |
Modellünkben a diszperzió sem költséggel, sem veszéllyel nem járt. Vajon befolyásolja-e általános következtetéseinket, ha a diszperzió költségeit is figyelembe vesszük? Tegyük fel, hogy a távolabb lerakott petéknek csupán az 1/3-a kel ki. Ekkor a mutáns rátermettsége 1+3·1/3 ·1/5 =1,2. Az elterjedt típusnak továbbra is vagy 1 saját utóda lesz, vagy esetenként 4/5 az esélye annak, hogy az õ petéje válik ivaréretté. Tehát a diszperzió elõnyeit ki nem használó típus átlagos utódszáma továbbra is kisebb lesz, mint egy, míg a mutáns rátermettsége határozottan nagyobb, mint egy. Azaz még ilyen jelentõs diszperziós veszteség esetén is megéri vándorolni (2.b ábra)!
Természetesen, ahogyan a mutáns terjed, úgy az elõnye egyre csökken, hiszen egyre többen lesznek a populációban azok, akik az õ fegyverével élve távoli petézõhelyeken is elhelyeznek utódokat. Pontos matematikai számításokkal kimutatható, hogy létezik egy ún. evolúciósan stabil diszperziós szint, az ettõl eltérõ szintû mutánsok kisebb rátermettségûek lesznek, tehát a szelekció ezt a diszperziós rátát tartja fenn.
Csupán az okoskodás leegyszerûsítése kedvéért határoztuk el, hogy a nõstény szûznemzéssel szaporodik. A fentihez hasonló számolás elvégezhetõ szexuális populációk esetén is, és megmutatható, hogy a nõstényeknek továbbra is megéri elvándorolni, még akkor is, ha az összes hím új helyre vándorol. A különbség csupán annyi, hogy az evolúciósan stabil diszperziós szint kisebb lesz, mint szûznemzéskor, s a nõstények csak akkor fognak vándorolni, ha a diszperzió nem túlzottan veszélyes.
A szexuális szaporodás figyelembevételével a diszperzió evolúcióját egy új, eddig nem említett módon is magyarázhatjuk. A szexuális szaporodás következménye, hogy a szülõk genetikai állománya keveredik az utódban. Ez csak akkor jelenthet elõnyt, ha a szülõk genetikai állománya különbözõ. Kicsi szaporodóközösségekben viszont a genetikai sokféleség gyorsan csökken, a populáció beltenyésztett lesz. A beltenyésztett populációkban az egyedek rátermettsége ahogyan azt a nemesítõk már több száz éve felismerték csökken. Tehát a szexuális populációkban az egyedek kellõ keveredése, azaz diszperziója növelni fogja az egyedek rátermettségét. Ezzel nagyon sok szexuális faj diszperziós viselkedése magyarázható, de az, hogy például az emlõsöknél miért a hímek, a madaraknál miért inkább a nõstények a vándorlók, továbbra sem érthetõ világosan. A következõ fejezetben a feltételes diszperzió meghatározása után erre a kérdésre is választ kapunk.
Feltételes diszperzió
A magasabb rendû fajok egyedei életmenetüket nagyon finoman képesek szabályozni. Diszperziós stratégiájuk függhet az egyed nemétõl, életkorától, egészségi állapotától, az élõhely minõségétõl vagy a populáció denzitásától. Az ilyen diszperziós stratégiákat összefoglalva feltételes diszperziónak nevezzük.
Ezek után térjünk vissza az elõzõ fejezet végén megfogalmazott kérdésre: a madaraknálmiért a nõstények diszperziója a jellemzõ, az emlõsöknél pedig miért a hímeké? Azaz, miért függ a szétszóródási hajlam az egyed nemétõl? Pusztán a beltenyésztés elkerülése elérhetõ bármilyen, a nemektõl független vándorlási szokás kialakításával. Mivel a madarak többsége monogám, az emlõsök viszont általában poligám életmódot folytatnak, kézenfekvõnek látszik, hogy a nemektõl függõ diszperziós stratégiák szoros kapcsolatban vannak a párosodási rendszerek ezen tulajdonságaival, így a legvalószínûbb magyarázat a következõ: az a nem fog helyben maradni, amelynél a diszperzió nagyobb késést (s így nagyobb veszteséget) okozna a párosodásban. A poligám rendszerekben a hímek között rendkívül erõs versengés folyik. Ezért a fiatal, tapasztalatlan és gyenge hímek nem tudnak párosodni. Ha vándorolnak, azzal nem veszíthetnek semmit, esetenként viszont szabad nõstényre találhatnak. A fiatal ivarérett nõstények ezzel szemben biztos, hogy párosodni fognak. ket csak késleltetné a párosodásban, ha vándorolnának. A monogám fajoknál a hímek általában értékes territóriumokat õriznek, annak elhagyása komoly veszteséget jelent. A nõstényeknek viszont érdemes a territóriumokat fölkeresni, és válogatni a tulajdonosok között.
A poligám fajoknál tehát nemcsak az egyed nemétõl, hanem annak életkorától is függött, hogy elõnyös-e számára diszperzió vagy sem. Ha a fiatal, élete teljében lévõ szülõ közvetlen környezetében az utódnak nincs esélye az életben maradásra vagy a szaporodásra, akkor mind a szülõnek, mind az utódnak elõnyös, ha messzire kerülnek az utódok. Például egy zárt erdõben élõ hatalmas kõrisfa közvetlen közelében nem tudnak a magok kikelni, mert nincs elég fény a számukra. A kõrisnek érdemes tehát messzire eljuttatnia a magokat. Ezt a növény a mag köré épített röpítõkészülék segítségével meg is teszi. Ahogy a fa öregszik, egyre nagyobb az esélye annak, hogy egy vihar vagy betegség elpusztítja. Halála után, az erdõ lombkoronáján keletkezett résen beáramló fény a talajban szunnyadó magok tucatjait ösztönzi arra, hogy csírázásnak induljanak. Kõrisünknek tehát elõnyös, ha idõsebb korában egyre több magot hullat saját maga közvetlen közelébe, hiszen így növekszik az esélye annak, hogy a helyét késõbb saját utóda fogja elfoglalni. Ophelie Ronce, francia kutatónõ és munkatársai kimutatták, hogy a különbözõ életkorú virágoskõris-populációk egyedei (Fraxinus ornus) más diszperziós stratégiát követnek. Ugyanez a kutatócsoport az elevenszülõ gyíknál (Lacerta vivipara) egyértelmûen bizonyítani tudta, hogy a diszperzió mértéke függ az életkortól. Ez a gyík, mely az Alföld õslápmaradványain hazánkban is megtalálható, egész Eurázsiában elterjedt. A kutatók öt éven keresztül tanulmányoztak egy dél-franciaországi populációt. Mesterségesen betelepített fiatal egyedek sorsának követésével kimutatták, hogy a megnövekedett populációméret növeli a halálozási rátát. Tehát a fiatalok a szülõkkel erõs versenyben állnak. Ezért a fiatalok jelentõs része szülõhelyétõl messzebb próbál meg boldogulni. Méréseik alapján egyértelmûvé vált, hogy a felnõtt nõstények túlélési esélye négyéves kor fölött gyorsan csökkenni kezd. Mindemellett a szülõ öregedésével nem változott az utódok fizikai állapota, ezzel szemben az idõsebb szülõk gyermekei jobban elviselték az idegen nõstényt maguk körül. Ekkor már sejthetõ volt, hogy a fiatal és az idõs anyák utódai kölönbözõ intenzitással fognak új élõhelyet keresni. Ezek után a terhes nõstényeket a várható szülés elõtt néhány nappal a laborba szállították, s az újszülötteket megjelölték. Néhány nap múlva mind az anyát, mind az utódokat visszahelyezték eredeti helyükre. Így az anya életkorát ismerve és a jelölt utódok sorsát követve mérni lehetett az utódok diszperziós stratégiáját az anya életkorának függvényében. Egyértelmûen bebizonyosodott, hogy a négyéves és annál idõsebb anyák nõstény utódainak vándorlási hajlama lényegesen kisebb, mint a fiatal anyák utódaié. A hím utódok (valószínûleg a beltenyésztési leromlás elkerülése érdekében) továbbra is az anya életkorától független vándorlási stratégiát követnek (3. ábra).
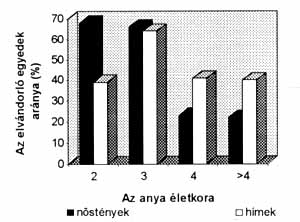
3. ábra. Az elevenszülô gyíkutódok
diszperziója az anya életkorának függvényében.
A nôstény utódok lényegesen kisebb arányban
vándorolnak új élôhelyre, ha az anya négyéves,
vagy annál idôsebb, a hím utódoknak a vándorlási
hajlama független az anya életkorától. (Ronce
et al. 1998 nyomán)
Természetesen a térben és idõben nem állandó veszélyt jelentõ élõsködõk vagy ragadozók veszélyességük miatt is feltételes diszperziós viselkedéseket alakíthatnak ki. Különbözõ tetûfajoknál bizonyított, hogy a ragadozók gyarapodásával növekszik az elvándorlók aránya is. Egy csüllõpopuláción (Rissa trydactyla) végzett mérés alapján a madarak elvándorlásának mértéke arányos a helyi élõsködõk okozta rátermettség csökkenésével. Nyilván a ragadozó vagy az élõsködõ is a körülményekhez alkalmazkodó diszperziós stratégiákat alakít ki. Ezért a szoros ökológiai kapcsolatban lévõ fajoknál a diszperziós stratégiák együttes, kölcsönható evolúciója, szakkifejezéssel koevolúciója játszódik le.
Az elõzõ fejezet elején láthattuk, hogy a térben és idõben heterogén környezetben elõnyös a feltétlen diszperziós stratégia. Azonban még több utóda lesz annak a típusnak, amely ismerve az élõhelyet valamilyen élõhelyfüggõ diszperziós stratégiát alakít ki. Az 1. ábrán bemutatott példánknál maradva, a legjobban az a típus jár, amelyik jó évben az I. foltban, rossz évben a II. foltban tartózkodik. Így átlagosan Ö16=4 utóda lesz évenként, ami az adott körülmények között a legjobb megoldás.A jól tájékozódó fejlett memóriával rendelkezõ állatok képesek ilyen stratégiák kialakítására.
A valóságban az élõhelyek eltartóképessége véges, ezért a populáció denzitásának növekedésével a szaporodóképesség csökkenni (vagy a halálozási ráta növekedni) fog. Ha az élõhely stabil, ennek eredményeként gyakran kimutatható, hogy a populáció mérete is egy stabil érték közelében marad. Ez azt jelenti, hogy mind az I., mind a II. foltban a nettó növekedési ütem nulla. Tételezzük fel, hogy az I. folt ezer, a második folt száz egyedet képes eltartani. Vajon, egy kellõen kifinomult diszperziós stratégia elterjedhet-e ebben a populációban? Bizonyos matematikai módszerek segítségével megmutatható, hogy egy térben heterogén, de idõben állandó környezetben az a diszperziós stratégia lesz evolúciósan stabil, melynek erõssége fordítottan arányos az adott élõhely eltartóképességével. Példánkban ez azt jelenti, hogy ilyenkor az I. foltban lévõ egyedek vándorlási kedve a tizede a II. foltban tapasztalható vándorlási szintnek. Tehát az evolúciósan stabil diszperziós stratégia függ az adott folt minõségétõl.
A vázolt gondolatnak egyenes következménye, hogy egy fokozatosan növekedõ populációnál kezdetben nem érdemes elvándorolni, ám ahogy az élõhelyen az egyedek száma és (vagy) azok mérete növekszik, úgy egyre kívánatosabb új területeket meghódítani. Ilyen denzitásfüggõ diszperziós stratégiát bizonyítottan sok faj alkalmaz. Az írás elején láttuk, hogy a térben és idõben heterogén környezetben a feltétlen diszperzió elõnyös. Gondolom nem meglepõ, hogy a denzitásfüggõ diszperzió ezeken az élõhelyeken még nagyobb rátermettségnövekedést okoz, ezért érthetõ, miért találkozunk vele gyakran az élõvilágban. Például az úgynevezett szubszociális pókok diszperziója is denzitásfüggõ. Ezekre a fajokra jellemzõ, hogy a petékbõl kikelt fiatal egyedek még egy ideig együtt maradnak az anyával. (A szociális fajoknál az egyedek egész életükben együtt maradnak.) Közismert, hogy a nagy eretnekpók (Amaurobius ferox) fiatal egyedei aktívan együttmûködnek a zsákmány elfogásában. Azonban ahogy a pókok növekednek, az anya hálója egyre kevésbé tud elegendõ zsákmányt szolgáltatni a számukra. Ennek hatására hirtelen intenzív elvándorlás kezdõdik, s tíz-húsz nap alatt mindenki elhagyja szülõhelyét. Azt is tudjuk, hogy elõször a kisebb testméretû pókok hagyják el a hálót. Nyilván ezek azok, melyek a nagyobb és agresszívebb testvéreik mellett nem tudnak elegendõ táplálékhoz jutni.
Összegzés, kitekintés
A térben és idõben változó környezetben a feltétlen diszperzió növeli a rátermettséget. Azt is megmutattuk, hogy a diszperzió elõnyös akkor is, ha a környezet stabil, de az utódok között nagyon erõs a verseny, vagy a beltenyésztés okozna csökkenést a rátermettségben. Azok az élõlények, amelyek képesek a külsõ körülmények figyelembevételével kialakítani diszperziós stratégiájukat (feltételes diszperzió), még hatékonyabban tudnak alkalmazkodni élettelen és élõ környezetükhöz. Ilyen a nemektõl függõ diszperzió is. Egy kísérlet bemutatásával igazoltuk, hogy az életkorfüggõ diszperzió nem csak az elméleti biológusok agyában létezõ stratégia. Végül a denzitásfüggõ diszperzió elõnyeirõl tettünk említést.
Összegezve megállapíthatjuk, hogy számos különbözõ mechanizmus képes tér- és (vagy) idõbeli heterogenitást létrehozni. A külsõ élettelen környezet sokfélesége kézenfekvõ, de a populációban meglévõ belsõ sokféleség (életkor és erõnlét sokfélesége, a nemek léte stb.) is ide sorolható. A diszperzió, különösen a feltételes diszperzió az egyedek egy olyan összetett tulajdonsága, mely hordozójának elõnyt jelent, ha az élõ és (vagy) élettelen környezete valamilyen heterogenitással rendelkezik.
A kutatóknak komoly kihívást jelent, hogy több, egymástól független, ráadásul egymást nem szükségképpen kizáró mechanizmussal magyarázható egy-egy faj diszperziós aktivitása. A diszperziót egyszerre tarthatja fenn például a táplálékért vagy a partnerért való versengés, és ezek mellett a beltenyésztési leromlás elkerülése, esetleg az idõnként felbukkanó erõs élõsködõveszély. Sajnos, mint az ökológia és az evolúcióbiológia oly sok más területén, kísérleteket végezni itt is nagyon nehéz, idõigényes és költséges feladat. Egyetlen olyan kísérlet ismert, ahol a diszperzió evolúcióját sikerült közvetlenül nyomon követni. A kísérletet a laboratóriumok kedvencén, az Echerichia coli baktériumon végezték japán kutatók. Ha tápoldattal feltöltött fõzõpoharakba E. coli bacikat szaporítunk, akkor egyes baktériumtörzsek következetesen az edény falára tapadnak (fal+), mások szabadon lebegnek az oldatban (falo). Tehát a fal+ a helyhez kötött, a falo a diszpergáló stratégiának tekinthetõ. Nakajima és mts. észrevették, hogy a baktériumok eme tulajdonsága öröklõdik. Ezek után a fõzõpoharakban többféle élõhelyet alakítottak ki. Az egyik esetben a fal+ típusú baktériumokat szaporítottak a pohárban, majd egy idõ múlva mintát véve a populációt átoltották egy új tápoldatba. Ezt a mûveletet sokszor elvégezték. Nyilvánvaló, hogy az átoltáskor azok a típusok jutnak el a következõ tápoldatba, melyek kevésbé tapadnak a pohár falára. Valóban ez is történt: fal+ baktériumok között rövid idõn belül megjelent a falo mutáns, majd néhány tucat átoltás után kiszorította a fal+ típust. A fordított kísérletet is elvégezték: a baktériumtenyészetbe eleinte falo típusokat oltottak, majd meghatározott idõnként az oldatot óvatosan leszívták, és helyére friss tápoldatot öntöttek. Így a pohárban a falra tapadók maradtak nagyobb arányban. A kísérlet a várakozásnak megfelelõen alakult, a fal+ felszaporodott a populációban, a falo típus pedig eltûnt.
E nagyszerû kísérlet ellenére a cikkben felsorolt
megállapítások többsége pusztán
a matematikai modellekbõl levont következtetéseken alapszik.
Azonban az utóbbi években érezhetõ, hogy a
kísérleti és terepi szakemberek egyre jobban törekednek
az elméleti jóslatok tesztelésére, így
várhatóan néhány éven belül a diszperzó
evolúcióját magyarázó hipotézisek
is biztosabb alapokra helyezhetõk.
Irodalom
Hamilton, W. D. and May, R. M. Nature 269: 578581, 1977
Jánosi, I. I. and Scheuring, I. J. of Theor. Biol. 187:
397408, 1997
Johnson, L. M., and Gaines, M. S. Annu. Rev. Ecol. Syst. 21:
449480, 1990
Kim, K. W. Behav. Ecol. Sociobiol. 48: 182187, 2000
McPeek, M. A. and Holt, R. D. Am. Nat. 140: 10101027, 1992
Nakajima, T. and Kurihara, Y. Oikos 69: 217223, 1994
Ronce, O., Clobert, J. and Massot, M. Proc. Natl. Acad. Sci. USA
95: 600605, 1998
| Természet Világa, | 132. évfolyam, 2. szám, 2001. február
https://www.kfki.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez