Könczöl Katalin
Premontrei Szent Norbert Gimnázium, Gödöllő
Fénynek nevezzük az elektromágneses hullámok 200–10 000 nanométerig (1 nm=10–6 m) terjedő tartományát. Az emberi szem ennek csak a töredékét képes érzékelni, körülbelül a 400–800 nm hosszúságú hullámokat. Az élőlények többségének fontos a fény érzékelése. A növényeknek a fotoszintézishez, az állatoknak a tájékozódáshoz fontos. Természetesen az egysejtű és a bonyolultabb testfelépítésű állatok is másként, és mást érzékelnek a körülöttük lévő világból.
A fény érzékelésének az első lépése a fény és a sötétség megkülönböztetése. A második lépcsőfok az, amikor az élőlény már a fény intenzitását és a fényforrás irányát is meg tudja határozni. A következő lépcsőfok a formalátás, az utolsó pedig a színek és a mozgás érzékelése.
Az egysejtűek a sejthártyájukkal érzékelik a fény intenzitását, és ennek változására valamilyen mozgással válaszolnak. Az ostorosmoszatoknál már szemfoltot is találhatunk. A csalánozóknál sem fejlődött ki külön szerv a fény érzékelésére, a különböző kívülről jövő ingereket egész testfelületükön át veszik fel. Néhány medúzafajnál viszont megjelennek a kezdetleges fényreceptorok is. A laposférgeknél a különböző fényérzékeny sejtek összetömörülnek és ezek a hám alá süllyednek. Így kezdetleges csésze- és gödörszemek alakulhatnak ki. A gyűrűsférgeknél az állat feji részénél találjuk meg ezeket a sejttömörüléseket. Egyes fajoknál már találkozhatunk bonyolult felépítésű látószervvel (pl. lencseszemmel) is. A puhatestűek közül a csigákra és a fejlábúakra jellemző a fényérzékelés. A kagylóknál csak néhány fajnál van „szem” a köpeny szélén. A csigák két pár tapogatója közül a hosszabbak végén „ülnek” a gödörszemek. A fejlábúaknak már kép- és színlátásra alkalmas hólyagszemük van, mely a hám betüremkedéséből alakult ki.
Az ízeltlábúak szeme a pontszem vagy az összetett (facetta-) szem, amely több száz egyszerű szemből áll. A rákoknál a facettaszem mellett találunk fényérzékeny dúcidegsejteket is. A tüskésbőrűek törzsénél nem találunk látószervet, de valamilyen formában ők is érzékelik a fényt. Ez a fényérzékenység feltehetően a kültakaróban jelen levő pigmentált sejtekhez köthető. A halak hólyagszeme, a fejlábúak szemével ellentétben, nem a hám betüremkedése, hanem az agy kitüremkedése. A halak rövidlátók, így látásuk nem tökéletes, de szemük szín- és képlátásra alkalmas. A kétéltűek látószerve igen fejlett, de csak a mozgást érzékelik. A hüllők, a madarak és az emlősök látószerve is jól „szervezett”. A kígyók két szemhéja átlátszó és összenőtt, ezért nem pislognak. Egyes madarakban a hipotalamusz bizonyos idegsejtjei érzékelik a koponyatetőn átszűrődő gyenge fényt. Így érzékelik a kakasok is a hajnal közeledését.
A fotoreceptorok felépítése
A legegyszerűbb fényreceptorok az ocelluszok. Szerkezetük szerint lehetnek pigmentált hámsejtek tömörülései vagy a hámsejtek betüremkedései révén létrejövő serleg vagy bogyó alakú bonyolultabb szemek is. Ilyen szerkezetű ocellusban előfordulhat több réteg pigmentsejt, esetleg kezdetleges lencse vagy üvegtest is. Ezek a behatoló fényhez viszonyítva inverz vagy everz szemek lehetnek. Az előbbi esetben a fénynek több közegen (lencsén, üvegtesten stb.) kell áthaladnia ahhoz, hogy elérje a fényérzékeny sejteket. Az utóbbiaknál a fény közvetlenül a receptorsejtekre vetődik.

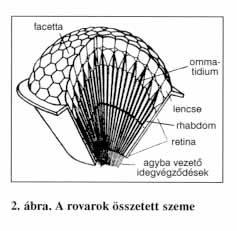
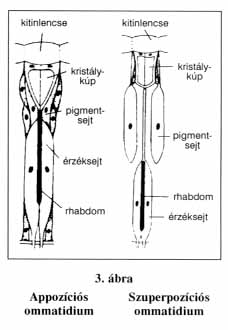
A főként rovaroknál előforduló összetett szemek ennél bonyolultabb felépítésűek. A facettaszem akár 12 000 egyszerű szemből (ommatidiumból) is állhat. Az ommatidiumnak két fő része van. A felszínen helyezkedik el a fénytörő apparátus (lencse), míg alatta az érzékelő idegsejtek rétege. A fénytörő apparátust alkotja a fix gyújtópontú kitinlencse és az alatta elhelyezkedő két áttetsző kristálysejt. Ez alatt találhatók a fényérzékeny retinulasejtek. Minden ommatidium nyolc retinulasejtet tartalmaz, melyek összessége alkotja a retinát. Minden retinulasejt elsődleges érzéksejt, melyek belső felszínén vékony mikrobolyhokból álló fényérzékeny szegély található, a rhabdom. Az egy ommatidiumhoz tartozó rhabdomok összessége adja az ommatidium fényfelvevő készülékét. Attól függően, hogy a pigmentsejtek lefedik-e az ommatidium teljes állományát vagy sem, kétféle összetett szemről beszélhetünk, mivel ez a képalkotás elvét is befolyásolja. Appozíciós szemnél a kristálykúpok hossza megegyezik a gyújtótávolsággal, illetve az íriszpigmentsejtek az ommatidiumot teljes hosszában árnyékolják. Ekkor a fénynyalábok egyenként jutnak be az ommatidiumok területére. Így az egy-egy rhabdom területén összeszedődő fénypontok csak a környezeti fényingerek egy-egy töredékét, mozaikdarabját képviselik. Az ommatidiumok összességének retinális részimpulzusaiból alakulhat ki a környezet valamelyest teljes képe. Az ilyen appozíciós szem a nappali életmódú rovarokra jellemző. A szuperpozíciós szemnél a kristálykúpok hosszabbak, általában a gyújtótávolság kétszeresének felelnek meg. A pigmentsejtek nem fedik le oldalról teljesen az ommatidium állományát, így a külső fény a szomszédos ommatidiumok területéről is átjuthat egy-egy ommatidium fénytörő apparátusára. Ily módon egy bizonyos számú kristálykúp egységes fénytörő közeget alkot, ami lehetővé teszi a gerincesek szeméhez hasonló összetett kép alkotását. Ez a szemtípus az éjszakai rovarokra jellemző, mivel kis fényintenzitás esetén is hatásosan összegyűjti a fényingereket.
A gerincesek látószervét a szemgolyó és a hozzá tartozó járulékos
szervek (pl. szemhéjak, szempillák) alkotják. Mivel az emberi szem felépítése
az összes többi gerinces állat szeméhez hasonló, a gerincesek szemének
a felépítését az emberi szemen keresztül mutatom be.
 |
 |
Az emberi szem három rétegből épül fel. A külső réteg ínhártyából és a szaruhártyából (cornea), a középső az érhártyából, a sugártestből és a szivárványhártyából, a belső réteg pedig az ideghártyából (retina), az ingerfelvevő rétegből áll. Mindhárom réteget „átfúrja” a látóideg. Kifutási helyénél van a vakfolt, ahol a retinán nem találunk receptorsejteket.
A pupillát körülölelő szivárványhártya mögött a szemlencse, a szemlencse és az ideghártya között pedig a kocsonyás üvegtest található. Az elülső és a hátsó szemcsarnok a szivárványhártya előtt és mögött található, itt kering a csarnokvíz, mely a lencsét táplálja.
Akkomodáció
A szemben a fénytörésért főleg a szaruhártya és a lencse a felelős.
A szem fénytörő képességét dioptriában (D) adjuk meg. A szaruhártya fénytörő
képessége minden pontján azonos, míg a lencsénél ez nincs így. Attól függően
változik, hogy a lencse magját vagy réteges köpenyét vizsgáljuk. Ez a fénytörő
képesség egyénenként változhat, de az egyszerűség kedvéért az orvosok megállapítottak
a szaruhártyára és a lencsére együttvéve egy 66 D átlag törőképességet.
A szem alkalmazkodását (akkomodációját) a lencse és a szem izmai
teszik lehetővé. Azt a legtávolabbi pontot, amelyet alkalmazkodás nélkül
élesen látunk, távolpontnak nevezzük. Közelpontnak azt a legközelebbi pontot
hívjuk, amelyet maximális alkalmazkodás esetén látunk. A közelpont fiatal
korban egészséges szem esetén 10 cm távolságban, a távolpont a végtelenben
van. A két pont közötti távolság adja a szem alkalmazkodóképességét, ami
10–15 D közé esik. A korral a lencse és a lencsefüggesztő rostok is vizet
veszítenek, így megváltozik a lencse alkalmazkodóképessége. Emiatt van
szüksége az idősebb embereknek „olvasószemüvegre”.
 |
 |
A szem akkomodációja természetesen együtt jár a pupilla nyílásának változásával. Ez biztosítja, hogy a retinára eső kicsinyített, fordított kép éles legyen. Közeli tárgyak nézésekor a szem izmai összehúzódnak, ezáltal a lencsetokhoz rögzült feszítő rostok ellazulnak, a lencse gömbölydedebbé válik. Ekkor a pupillák összeszűkülnek. Távoli tárgyak nézésekor ennek a fordítottja játszódik le. (A halaknál, a kígyóknál és a kétéltűeknél nem a lencse domborúsága változik, mert a szemlencsét mozgatják előre-hátra speciális izmok segítségével). A pupilláknak nem csak ez az alkalmazkodása ismert. A szembogár akkor is összeszűkül, ha világítás éri. Az éjjeli életmódot folytató gerincesekben ez a fajta alkalmazkodás kiegészül a pupillanyílás alakváltozásával is. Ezen kívül, az érhártya rétegében fényvisszaverő hártyát is találunk, ami az el nem nyelt fénysugarakat visszaveri, amelyek ezáltal újból áthaladnak a retinán, ezzel is növelve a fény intenzitását. Ez okozza például a macska vagy a gazella „fénylő” szemét is.
Pálcikák és csapok
A retina, vagyis az ingereket felfogó réteg, három sejtrétegből áll. A csapok és pálcikák rétegéből, a bipoláris sejtek rétegéből, ahol amakrin és horizontális sejteket is találunk, illetve a ganglionsejtek rétegéből.
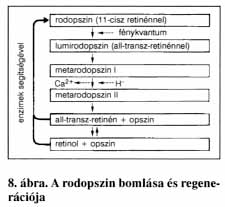
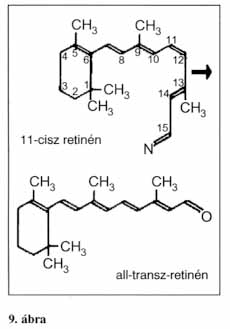
A fényérzékeny sejtek a retina hátulsó oldalán alkotnak hálózatot (inverz szem). A pupillával szemben találjuk az éleslátás helyét, a sárgafoltot, amelyet csak csapok alkotnak. A retina széleihez közeledve egyre több pálcika vegyül a csapok közé. Ezek külső szegmense (kültagja) nyeli el a fényt, majd biokémiai folyamatok során idegi jelekké változtatja azt. Mindkét sejttípus kültagjában fényérzékeny membránrendszer található. A pálcikák külső szegmense henger alakú. A pálcikák gyenge fényben működnek, a sötétség–fény megkülönböztetése a feladatuk, az éjszakai éleslátásért felelősek. Ez összefügg a rodopszin nevű (opszinból és retinolból felépülő molekula) festékanyag bomlásával. A molekula fény hatására először opszinra (fehérje) és 11-cisz-retinénre (A-vitamin-származék) bomlik. A retinén azonnal transz-retinénné polimerizálódik. A bomlási folyamat úgynevezett metarodopszin–II stádiumában a molekulában Ca-ionok H-ionokra cserélődnek. A szabad Ca-ionok a membránhártyához kötődnek, ezzel gátolva a Na-ionok beáramlását a sejtbe. Ez potenciálváltozást okoz a sejtben, ami jelekként továbbítódik az agyba a pálcikák szinaptikus végződésein keresztül. Sötétben, enzimek segítségével a két molekula újból rodopszinná egyesül. Bámulatra méltó a leírt reakció érzékenysége. Elég 5-6 rodopszinmolekulának szétesnie ahhoz, hogy ez már potenciálváltozást okozzon a sejtben, melyben összesen 10 millió ilyen molekula található.

A csapok kültagja kúp alakúak. A csapok a nappali éleslátásért
és a színlátásért felelősek. A csapokban háromféle fényérzékeny festék
található. A csapok egy részében a zöld, másokban a piros, megint másokban
a kék fényre érzékeny pigmentek vannak. Ezek együttes vagy egyedi reakciója
teszi képessé az emberi szemet a közel 160 színárnyalatnak a megkülönböztetésére.
(Itt meg kell említeni, hogy a színlátást bizonyossággal csak a főemlősökben
igazolták.) Tehát a színérzékelés és a nappali látás a csapok, míg a szürkületi
látás lényegében a pálcikák tevékenysége. (A-vitamin hiánya esetén ezért
lép fel a szürkületi vagy farkasvakság.) A színvakság, illetve színtévesztés
a csapok teljes vagy részleges működésképtelensége miatt jön létre). Az
emberi szemben körülbelül 120 millió csap- és 6 millió pálcikasejt biztosítja
az éleslátást. Az éjszaka aktív élőlények szeméből teljesen vagy csak részlegesen
hiányozhatnak a csapok, mivel nekik nincs szükségük a színek érzékelésére.
Emiatt viszont szemük a félhomályban sokkal érzékenyebb.
 |
 |
 |
Fényérzékelés
A fényfelvevő réteg által kibocsátott elektromos ingerek több szinapszison keresztül jutnak el az agyig. A pálcikák és a csapok ezeket a jeleket először a bipoláris sejteknek, majd ezek a ganglionsejteknek továbbítják. A ganglionsejtek az agynak küldik tovább a csapok és pálcikák információit. Az, hogy a ganglionsejtek küldenek-e idegi jeleket az idegközpont felé, és ha igen, akkor milyeneket, az attól is függ, hogy az adott ganglionsejthez tartozó receptormező (RM, vagyis az egy ganglionsejthez tartozó receptorsejtek összessége) on vagy off központú-e.
Az on központú RM azt jelenti, hogy a ganglionsejt akkor küld jeleket az agynak, ha a RM-nek csak a közepe kap fényt. Az off központú RM pedig akkor küld jeleket, ha a RM közepére eső fény kialszik. (Az emberi szemben a ganglionsejtek ilyen megoszlása 50-50 százalékos.)
Tehát a ganglionsejtek a kontrasztokat érzékelik. Ezen alapszik az egyik legismertebb optikai csalódás, az úgynevezett Hermann-rács. A fehér csíkok kereszteződésében sötétebb foltok látszanak. A jelenség magyarázata a következő: Ha a rácsra nézünk, a rács képe a receptív mezőkre vetül. Az az on központú RM-hoz tartozó ganglionsejt, amelynek RM-je a csíkokra (de nem a kereszteződésre) esik, igen erős ingerületbe jön, hiszen a jórészt sötét háttér miatt gátlás nem alakul ki. A kereszteződésben viszont a gátló környezet kétszer annyi fényt kap, mint az előbb, az így gátolt idegsejt aktivitása lényegesen csökken. Ebből adódóan az agy azt az információt kapja, hogy a kereszteződések sötétebbek, mint a csíkok.
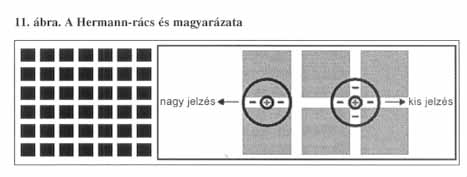
A ganglionsejtek főleg a megvilágítás kezdetét és végét jelzik az agynak. Tehát azt feltételezhetjük, hogy az agy képtelen a homogén (egyformán világos, vagy egyformán sötét) felületek, tárgyak észlelésére, kivéve ezek széleit. De a tárgy kontúrja is csak akkor vehető észre, ha a kép elmozdul a retinán az egyik ganglionsejt RM-jéből a szomszédos RM-re. Ha a tárgy nem mozog, akkor a szemünk teszi meg ezt helyette. A szemünk állandó, akaratunktól független mozgásban van. Tehát a retinának a tárgyhoz képest el kell mozdulnia ahhoz, hogy érzékelés jöhessen létre. Ha a retinára eső kép mozdulatlan, nem küldenek a ganglionsejtek impulzusokat az agy felé. (Ezért nem látjuk a retinánkra állandóan ugyanoda árnyékot vető ereket sem.) Így bármely tárgy nézésekor a szemgolyó mikromozgásokat végez. Ezért a mozgásért a hat pár szemizom a felelős.
Mozgásérzékelés
Most nézzük, hogyan érzékeljük a mozgást. Nyulakkal végzett kísérletekben vizsgálták az irányszelektív sejteket, a retinának azon sejtjeit, amelyek képesek felismerni a mozgás irányát. A nyulak retináját úgy ingerelték, hogy két rést egymás után megvilágítottak, ezzel mozgást szimulálva. A ganglionsejtek válaszjeleit a sejttesthez közel, de nem a sejten belül vizsgálták. Először egyesével világították meg a hasadékokat, majd egymás után (először balról jobbra, majd fordítva), ezzel mozgást érzékeltetve. A kutatók azt tapasztalták, hogy a ganglionsejt élénken reagál az egyik irányú mozgásnál, de a vele ellentétesre alig. Sőt, gyengébb válaszjelekkel válaszol, mintha csak az egyik rést világították volna meg. Ez meglepő volt, hisz ha a réseket egymás után világítják meg, akkor a receptorsejtek kétszer annyi fényt kapnak, mintha csak az egyik hasadékot világítanák meg. Ezekből azt a következtetést vonják le, hogy van egy „kitüntetett” irányú és egy „tilos” irányú mozgás. Ezek érzékelése pedig valamiféle szelektív gátlási folyamat során jön létre. Feltételezhető, hogy a fényérzékelő sejtek olyan elrendezésben kapcsolódnak az irányszelektív sejtekhez, hogy a „kitüntetett” irányú mozgás először a serkentő szerepet betöltő sejteket hozza működésbe, majd a gátló sejteket. A serkentő jelek gyorsabban terjednek, és előbb is keletkeztek a gátló jelekhez képest, így azok előbb elérik a ganglionsejteket, melyek ennek megfelelően válaszolnak. Így a gátló jel még gyengíteni se tudja a serkentő jel hatását. A „tilos” irányú mozgásnál először gátló jelek keletkeznek, s csak ez után, egy kicsit később a gyorsabb serkentő jelek. Ha beérik a lassabb gátló jeleket, kioltják egymást. Csendes gátlás jön létre akkor, ha a kétféle jel egyszerre érkezik meg a ganglionsejthez, így a serkentő jel gyengítetten, de hatást gyakorol a ganglionsejtre. Ezzel magyarázható a tiltott irányú mozgásra adott csekélyebb válaszreakció.

Érdekességek
Eddig az emberi szem számára is látható fénnyel foglalkoztunk, de sok állatfaj az ultraibolya és az infravörös tartományba eső hullámokat is képes érzékelni, mint a rovarok zöme. Ez adhat magyarázatot arra, hogy a méhek, annak ellenére beporozzák a pipacsot, hogy nem tudják érzékelni a piros színt, viszont a pipacs visszaveri az UV-sugarakat.
A kígyónak vannak olyan bemélyedései a fején, melyek segítségével érzékelni tudja az infravörös sugarakat. Így a kígyó még a sötétben is „lát”, vagyis érzékeli a környezeténél melegebb tárgyakat.

Bár a számunkra is érzékelhető fény tartományában lát a négyszemű hal, szeme nem szokványos felépítésű. A feje tetején lévő szemgolyói kettéosztottak egy vízszintes vonallal. Így nem okoz gondot a halnak a víz és a levegő határán keletkező fénytörés. Ha a víz színéhez közel lebeg, a szemei felső részével a levegőben repkedő rovarokat, míg szemei alsó felével a vízben élő ragadozókat kémleli. Mind a négy szemrészhez külön retina tartozik. A felsőre a vízből jövő, az alsóra a levegőből érkező fénysugarak vetődnek. Agya külön-külön értelmezi a vízből és a levegőből érkező ingereket.
A kaméleonnak csak egy pár szeme van, de ezek egymástól függetlenül mozgathatók.
A szerző Diákpályázatunkon a Varjú Dezső által kiírt a Biofizika, biokibernetika különdíj kategóriában II. díjat kapott.
Kisszótár
Amakrin sejtek: a bipoláris sejteket kötik össze a ganglion sejtekkel.
Bipoláris sejt: olyan idegsejt, melynek két egymással szembenálló nyúlványa van.
Dioptria: a lencsék fénytörő képességét adja meg; a méterben megadott fókusztávolság reciprok értéke.
Elsődleges érzéksejt: specializált idegsejt, néha elágazó ingerfelvevő nyúlványokkal.
Fókuszpont: az a pont, ahol a lencse a tengelyével párhuzamosan beeső fénysugarakat egyesíti.
Fókusztávolság: a fókuszpont távolsága a lencsétől.
Fotoreceptor: fényre érzékeny receptorsejtek.
Ganglion sejtek: dúc idegsejtek.
Hipotalamusz: az agy része, a szervezet létfenntartó működéseinek fő irányítója.
Horizontális sejtek: azok a sejtek, melyek a csapokat és a pálcikákat kapcsolják össze.
Pigmentált hámsejt: olyan kültakaróban elhelyezkedő sejt, mely festékanyagot tartalmaz.
Potenciál: egy pont elektromos tulajdonságainak a különbsége a nullához viszonyítva.
Receptorsejt: érzékelő sejt.
Szinapszis: két idegsejt közötti kapcsolat.
Irodalom
Günter Vogel – Harmut Angermann: SH atlasz, Biológia. Springer Hungarica,
Budapest, 1992.
Janusz Gill: Zarys fizjoloii porównawczej zwierząt.
Państwowe Wydawnictwo Naukowe, Warszawa,
1987.
Ádám György – Fehér Ottó: Élettan biológusoknak. Tankönyvkiadó, Budapest,
1991.
Jerzy Trojanowski: Biochemia dla biologów. Państwowe
Wydawnictwo Naukowe, Warszawa; 1974.
Erich Grosse: Z biologią za pan
brat, eksperymenty biologiczne, Państwowe
Wydawnictwo „Iskry”, Warszawa; 1976.
Dr. Donáth Tibor: Anatómiai atlasz. Medicina, Budapest, 1985.
Stéphane Frattini: Az élővilág rekordjai. Alexandra, Pécs, 1995.
Vigh H. Borbála – Kondics Lajos: Összehasonlító szövettan. Tankönyvkiadó,
Budapest, 1989.
Dr. Budó Ágoston: Kísérleti fizika. Tankönyvkiadó, Budapest, 1980.
https://www.mozaik.info.hu/mozaweb/Feny
| Természet Világa, | 133. évfolyam, 12. szám, 2002. december
https://www.chemonet.hu/TermVil/ https://www.kfki.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez